Содержание
как получить, где ее можно заказать, какие бывают выписки и зачем они нужны
Анастасия Корнилова
регулярно заказывает выписки из ЕГРН
Профиль автора
Без выписки из ЕГРН в России редко обходится хотя бы одна сделка.
Если хотите получить кредит под залог квартиры, взять с покупателя задаток при продаже объекта или подать иск против соседа, который установил забор на вашей территории, понадобится выписка из Единого государственного реестра недвижимости — ЕГРН. Это документ, который подтверждает права на квартиру, дом или другой объект недвижимости.
Единого срока действия у выписки из ЕГРН нет. Здесь ориентируются на требования ведомства или организации, которые ее требуют. Чаще всего выписка из ЕГРН действительна месяц. Но, например, поставить дом на охрану получится и с выпиской годичной давности.
Чтобы получить выписку, придется заплатить госпошлину. Сколько и куда — зависит от того, как подаете заявление — электронно или лично, в Росреестр или МФЦ. Все варианты, как ее заплатить, есть на сайте Росреестра.
Все варианты, как ее заплатить, есть на сайте Росреестра.
Что вы узнаете
- Какие бывают выписки
- Что такое выписка на объект недвижимости
- Что такое выписка из ЕГРН в отношении субъекта
- Как заказать электронную выписку в Росреестре
- Как заказать выписку через сайт Федеральной кадастровой палаты
- Как получить 100 выписок из ЕГРН за 460 Р
- Как заказать выписку через портал госуслуг
- Как заказать выписку в МФЦ
- Как заказать выписку на платных сайтах
- Кто может заказывать выписку и в каких случаях она требуется
Какие бывают выписки
Выписки из ЕГРН содержат информацию об основных характеристиках объекта, сведения о лицах, у которых есть права на объект, о кадастровой стоимости, обременениях и многом другом. Объем информации зависит от конкретного вида выписки и от того, кто запрашивает сведения. Например, если за выпиской обратится постороннее лицо, персональные данные правообладателя в выписке не укажут. Они будут видны только в выписке, которую получит сам собственник или его представитель по доверенности.
Они будут видны только в выписке, которую получит сам собственник или его представитель по доверенности.
В зависимости от сведений, которые нужны, можно получить два варианта выписок:
- В отношении объектов, то есть недвижимости.
- В отношении субъектов, то есть людей.
/guide/kak-kupit-kvartiru/
Как купить квартиру
Расскажу подробно про оба вида.
Что такое выписка на объект недвижимости
Это самые востребованные выписки. Выписку на объект может заказать любой человек или юрлицо, но надо знать точный адрес недвижимости, иначе ее просто не найдут в реестре.
Выписки в отношении объектов тоже бывают нескольких типов.
Выписка из ЕГРН об основных характеристиках и зарегистрированных правах на объект недвижимости. Она содержит подробную информацию о квартире, земельном участке, садовом домике или торговом центре, в том числе:
- Характеристики объекта, включая кадастровый номер, адрес, площадь, назначение, наименование, этажность, дату ввода объекта в эксплуатацию.

- Кадастровую стоимость объекта.
- ФИО правообладателя или название, если собственник — юридическое лицо.
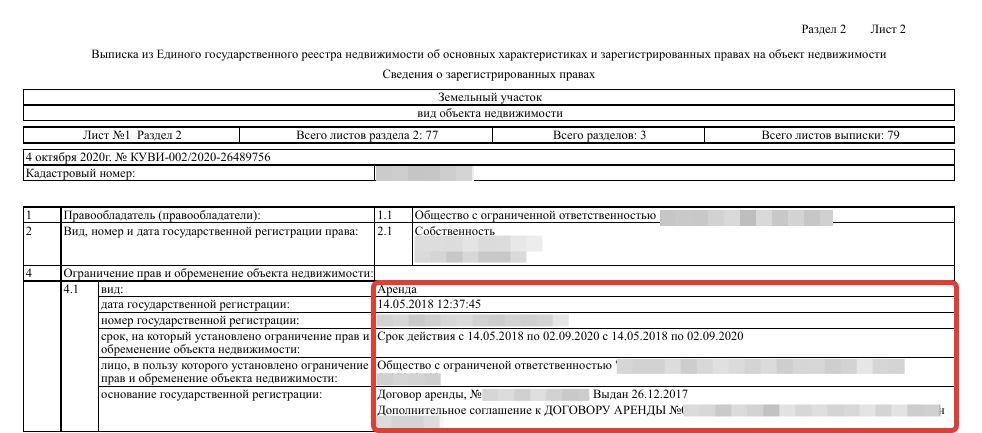
- Информацию об обременениях.
- План расположения помещений на этаже или схематическое отображение расположения объекта на земельном участке.

/kadastr-value/
Как узнать кадастровую стоимость недвижимости
Такая выписка стоит для граждан 390 Р на бумаге и 290 Р в электронном формате. Чаще всего ее заказывают, чтобы подтвердить свои права на недвижимость или проверить объект на обременения: например, узнать перед покупкой квартиры, нет ли на ней ареста. Если будете заказывать бумажный вариант через МФЦ, надо будет доплатить 70 Р. В этом случае выписка вам обойдется в 460 Р.
390 Р
стоит бумажная выписка из ЕГРН
Выписка из ЕГРН об объекте недвижимости. Ее еще называют расширенной выпиской. В ней, кроме сведений, которые есть в обычной выписке на объект, будут перечислены основания возникновения права. Например, что собственник приобрел квартиру на основании договора купли-продажи от определенной даты или что право собственности на земельный участок возникло по решению суда.
Например, что собственник приобрел квартиру на основании договора купли-продажи от определенной даты или что право собственности на земельный участок возникло по решению суда.
Еще там будут подробности об имеющихся обременениях, например ипотеке, наличии договора аренды, аресте или запрете на совершение сделок с объектом.
В расширенной выписке тоже будут сведения о координатах объекта, местоположении границ земельного участка, а также об адресах правообладателей смежных земельных участков. Если выписка заказана по земельному участку, в ней можно увидеть кадастровые номера объектов, которые расположены в границах этого участка.
740 Р
стоит расширенная выписка из ЕГРН
В этой же выписке будет указано, включен ли объект в реестр объектов культурного наследия, попадает ли земельный участок в границы охранной зоны или зоны с особыми условиями использования территории.
Еще в выписке указывают, например, информацию о невозможности регистрации без личного участия правообладателя или законного представителя.
Граждане могут получить такую выписку в бумажном виде за 740 Р, а в электронном — за 350 Р. При заказе бумажной выписки через МФЦ надо будет заплатить 870 Р.
Выписка из ЕГРН о переходе прав на объект показывает, сколько раз объект поменял собственника. В этом документе будут перечислены фамилии всех владельцев объекта. Если недвижимость слишком часто меняла владельцев, стоит проверить, не было ли по ней судебных споров.
/guide/concrete-docs/
Какие бывают документы-основания при сделках с недвижимостью
Информации о кадастровой стоимости объекта, залогах и обременениях в такой выписке не будет.
Гражданам такую выписку выдают за 290 Р, если документ электронный, и за 390 Р в бумажном варианте. Через МФЦ ее можно заказать за 460 Р — 70 Р возьмут еще за то, чтобы ее распечатать.
Выписка из ЕГРН о кадастровой стоимости объекта недвижимости позволяет проверить, правильно ли начисляются налоги. Такая выписка выдается бесплатно, и в ней указывают актуальные сведения о кадастровой стоимости недвижимости.
Такая выписка выдается бесплатно, и в ней указывают актуальные сведения о кадастровой стоимости недвижимости.
Что такое выписка из ЕГРН в отношении субъекта
Выписку из ЕГРН о недвижимости конкретного человека может получить только он сам. Единственное исключение — представитель собственника по нотариально заверенной доверенности.
Такая выписка пригодится в ситуации, если вы, например, продали квартиру, а налоговая продолжает слать уведомления об уплате налога. Или вы хотите получить жилье по договору социального найма, и вам надо подтвердить, что у вас в собственности нет жилой недвижимости.
Стоимость этой выписки зависит от объема запрошенных сведений. Если нужна информация о недвижимости в пределах региона, это будет стоить 640 Р в бумажном варианте и 470 Р в электронном виде. Если нужны сведения по всей России, выписка на бумажном носителе обойдется в 1770 Р, а в электронном варианте — 870 Р. Если обратитесь в МФЦ, надо будет заплатить 750 Р и 2080 Р соответственно.
/guide/doma-net/
Где взять справку об отсутствии жилья
Как заказать электронную выписку в Росреестре
Электронную выписку заказывают на сайте Росреестра. Она стоит 300 Р, а делают ее меньше трех дней. Заполнить форму можно быстрее, если воспользуетесь сервисом «Личный кабинет правообладателя». Авторизоваться в кабинете можно по учетной записи единого портала госуслуг. Но только если у вас подтвержденная учетная запись и электронная подпись.
На главной странице сайта выберите вкладку «Физическим лицам». Нажмите на вкладку «Получить сведения из ЕГРН» и перейдите к форме заявления Вход в личный кабинет находится справа в верхнем меню Без электронной подписи получить сведения через личный кабинет не удастся
Если нет электронной подписи и подтвержденной учетной записи или вы не хотите давать к ней доступ сайту Росреестра, можно заполнить форму и без авторизации. Это тоже рабочий способ, но заполнять придется больше полей.
По закону срок изготовления выписки — не более трех рабочих дней со дня получения запроса, но электронные выписки нередко приходят в тот же день.
/guide/e-signature/
Как получить электронную подпись
Проверьте, чтобы все поля, отмеченные красными знаками, были заполнены Пошагово заполните форму Вот так выглядит форма, если у вас есть электронная подпись. Если у вас ее нет — придется заполнять больше полей Проверьте все еще раз и отправьте запрос
Когда выписка будет готова, вы получите на адрес электронной почты, который указали в заявлении, ссылку на электронную выписку и ключ доступа.
Надо будет перейти по ссылке и заполнить необходимые поля. После этого загрузится архив с двумя файлами — выпиской и электронной подписью. Чтобы открыть файлы на своем устройстве, воспользуйтесь сервисом Росреестра. С его помощью можно сформировать выписку для печати и проверить подлинность электронной подписи.
Учтите, что собственник в любой момент может запросить сведения о лицах, которые интересовались его недвижимостью. Такая справка обойдется собственнику объекта в 390 Р за бумажный вариант и 290 Р за электронный. В справке будет информация обо всех, кто заказывал выписки из ЕГРН по недвижимости заявителя, будь то физлица, компании или органы власти. Единственное исключение — запросы от правоохранительных органов в рамках оперативно-разыскных мероприятий. Про них собственнику не скажут.
В справке будет информация обо всех, кто заказывал выписки из ЕГРН по недвижимости заявителя, будь то физлица, компании или органы власти. Единственное исключение — запросы от правоохранительных органов в рамках оперативно-разыскных мероприятий. Про них собственнику не скажут.
/prava/sobstvennik/
Права собственника квартиры
Как заказать выписку из ЕГРН через сайт Федеральной кадастровой палаты
Еще способ заказать выписку — воспользоваться сервисом, который есть на сайте Федеральной кадастровой палаты. Нужно авторизоваться через портал госуслуг, потом ввести кадастровый номер объекта, на который хотите получить выписку, и выбрать нужный вариант выписки.
Росреестр о сервисе ФКП для получения выписок
Выписка из ЕГРН об основных характеристиках и зарегистрированных правах на объект стоит 290 Р, об объекте недвижимости — 350 Р, о переходе прав — 290 Р.
Выписка будет готова уже через несколько минут. Прямо на сайте появится электронная папка — ее можно скачать на свой компьютер или планшет, открыть и получить готовый документ. Выписка будет подписана электронной подписью.
Выписка будет подписана электронной подписью.
Ссылка на сервис есть прямо на главной странице Нужно выбрать, какую выписку будете заказывать, и оплатить. Все будет готово через несколько минут
Как получить 100 выписок из ЕГРН за 460 Р
Если часто заказываете выписки, есть смысл воспользоваться более дешевым вариантом. Такой вариант удобен для предпринимателей, особенно для тех, кто работает с недвижимостью и кому нужно часто получать сведения о разных объектах. Людям, которым выписка нужна раз в год, а то и реже, вряд ли стоит пользоваться таким способом.
Срок действия оплаты Росреестр не указывает — вероятно, вы сможете пользоваться ключом, пока не выберете все оплаченное количество выписок. Но при условии, что законодательство не изменится. Чтобы оплатить и получить доступ к ФГИС ЕГРН, для начала нужно войти в личный кабинет на сайте Росреестра.
После входа вам будут доступны дополнительные опции. Выберите кнопку «Мои ключи» и нажмите «Сформировать ключ» Система сформирует ключ доступа. Скопируйте его куда-нибудь, чтобы потом быстро найти Теперь надо оплатить доступ. Это можно сделать через кнопку «Мой баланс». Выберите вкладку «Предоставление сведений ЕГРН» и нажмите «Внести оплату» Вы можете получить 100 выписок за 460 Р или 1000 за 1940 Р Оплачивать можно картой, со счета телефона и даже наличными через банк
Скопируйте его куда-нибудь, чтобы потом быстро найти Теперь надо оплатить доступ. Это можно сделать через кнопку «Мой баланс». Выберите вкладку «Предоставление сведений ЕГРН» и нажмите «Внести оплату» Вы можете получить 100 выписок за 460 Р или 1000 за 1940 Р Оплачивать можно картой, со счета телефона и даже наличными через банк
Когда деньги поступят, на странице «Мой баланс» на вкладке «Предоставление сведений ФГИС ЕГРН» появится количество доступных операций и количество объектов в пакете.
Теперь на странице «Запрос посредством доступа к ФГИС ЕГРН» можно ввести полученный уникальный ключ доступа и заказать выписку.
Просто скопируйте и вставьте ключ. Дальше будет стандартная процедура заказа выписки
Как заказать выписку через портал госуслуг
Через портал госуслуг можно заказать не все выписки.
В электронном виде доступны выписки на объект — обычная и расширенная и выписка о переходе прав Чтобы заказать выписку, нажмите на кнопку «Получить услугу» Ваши данные появятся в бланке автоматически, а сведения об объекте придется ввести вручную После подачи заявления вы получите сообщение с номером вашей заявки
Сервис не всегда срабатывает.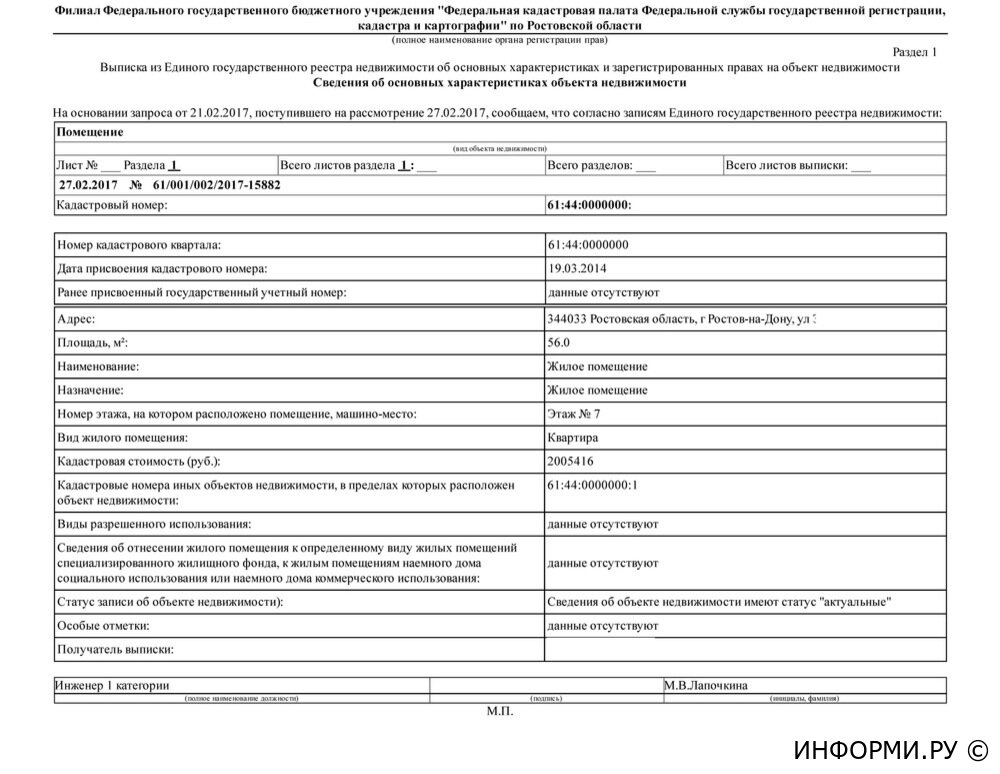 Я, например, не смогла получить выписку: дважды приходило уведомление про сбой в процессе обработки. Если вам удалось заказать и получить выписку через сервис госуслуг, напишите об этом в комментариях.
Я, например, не смогла получить выписку: дважды приходило уведомление про сбой в процессе обработки. Если вам удалось заказать и получить выписку через сервис госуслуг, напишите об этом в комментариях.
Еще на сайте госуслуг можно узнать, какие документы понадобятся в Росреестре, сколько будет стоить выписка, где и в каком виде ее можно получить.
Если хотите узнать, как получить сведения о большом количестве объектов, перейдите на вкладку «Предоставление сведений, содержащихся в ЕГРН, посредством обеспечения доступа к информационному ресурсу, содержащему сведения ЕГРН».
Еще можно узнать, как получить другие сведения, например кадастровый план территории В перечне сведений, которые можно получить в Росреестре, больше 30 позиций. Если нужна информация, как получить выписку из ЕГРН о правах отдельного лица на его объекты недвижимости, нажмите на соответствующую вкладку Допустим, нужен один регион, выписка в электронном формате. Это обойдется в 400 Р Есть пять вариантов, как заказать и получить документ В разделе «Контакты» будет адрес и часы работы отделения Росреестра в вашем городе
Как заказать выписку в МФЦ
В разделе «Контакты» на сайте госуслуг есть адреса многофункциональных центров. Чтобы заказать выписку, надо прийти в МФЦ с паспортом. Будьте готовы назвать оператору адрес нужного объекта или его кадастровый номер, а лучше и то и другое. Если хотите получить выписку в отношении человека, у вас должна быть от него доверенность с соответствующими полномочиями.
Чтобы заказать выписку, надо прийти в МФЦ с паспортом. Будьте готовы назвать оператору адрес нужного объекта или его кадастровый номер, а лучше и то и другое. Если хотите получить выписку в отношении человека, у вас должна быть от него доверенность с соответствующими полномочиями.
Квитанцию на уплату госпошлины можно оплатить заранее или взять бланк в офисе МФЦ и оплатить в ближайшей кассе любого банка. Посмотреть реквизиты можно на сайте Росреестра.
/guide/homebook/
Как получить выписку из домовой книги
В МФЦ надо взять талон на получение услуги и дождаться, когда пригласят. Сотрудник МФЦ сам заполнит и распечатает бланк заявления. Останется только все проверить и подписать. Как правило, выписку делают 3—5 рабочих дней с момента подачи заявления.
На бумажной выписке должна быть печать Росреестра или МФЦ и подпись ответственного специалиста или регистратора
Как заказать выписку на платных сайтах
В интернете много сайтов, которые предлагают заказать выписку из ЕГРН дешевле, чем в Росреестре. Такие сайты покупают информацию из реестра, а потом перепродают сведения.
Такие сайты покупают информацию из реестра, а потом перепродают сведения.
Но база данных на стороннем сайте может быть неактуальной. Еще среди этих сайтов попадаются откровенно мошеннические: они просто собирают деньги, ничего не предоставляя.
/real-estate/
Какие документы проверять перед покупкой квартиры
Я не рекомендую обращаться за выпиской на сторонние ресурсы. Пытаясь сэкономить 150—200 Р, рискуете получить устаревшие сведения или просто потерять деньги.
Кто может заказывать выписку и в каких случаях она требуется
Есть выписки, которые может получить только собственник, либо его законный представитель или представитель по доверенности, либо правоохранительные органы, суды, приставы, нотариусы и органы власти. Расскажу о таких выписках:
- Выписка из ЕГРН о признании правообладателя недееспособным или ограниченно дееспособным — может потребоваться судам и нотариусам, чтобы удостовериться в дееспособности человека, потому что, например, сделка с недееспособным правообладателем будет считаться недействительной.
- Выписка из ЕГРН о правах отдельного лица на имевшиеся (имеющиеся) у него объекты недвижимости.
- Выписка о лицах, получивших сведения об объекте недвижимого имущества.
- Выписка о содержании правоустанавливающих документов — может понадобиться, если утрачены оригиналы документов на недвижимость. Из этой выписки можно получить информацию, на основании чего право появилось и на каком основании сохраняется.

Остальные выписки может заказать любой человек. Но прежде чем заказывать, уточните, какая именно выписка и в каком варианте нужна.
/netudoma/
Какие документы проверить при аренде квартиры
Если никаких особых требований к содержанию и форме нет, проще получить электронную — не придется ехать за бумажным экземпляром, можно заказать и получить выписку, не выходя из дома или офиса.
Какие выписки обычно требуются и в каких случаях
| Название | Для чего |
|---|---|
| Выписка об основных характеристиках и зарегистрированных правах | Подтвердить потенциальному покупателю право собственности на недвижимость Зарегистрироваться по месту пребывания Вступить в наследство |
| Выписка из ЕГРН о переходе прав на объект | Для банка при ипотечной сделке При покупке квартиры, чтобы посмотреть историю перехода прав собственности на квартиру |
| Выписка из ЕГРН о правах отдельного лица на объекты недвижимости | Для налоговой, чтобы подтвердить, что квартиры или дома у вас больше нет Для органа, который принимает решение о предоставлении жилья Для банка при решении вопроса о большом кредите |
| Выписка о кадастровой стоимости | Для налоговой, чтобы проверить правильность начислений |
Выписка об основных характеристиках и зарегистрированных правах требуется, чтобы:
- Подтвердить потенциальному покупателю право собственности на недвижимость.
- Зарегистрироваться по месту пребывания.
- Вступить в наследство.

Выписка из ЕГРН о переходе прав на объект нужна:
- Банку при ипотечной сделке.
- При покупке квартиры, чтобы посмотреть историю перехода прав собственности на квартиру.
Выписка из ЕГРН о правах отдельного лица на объекты недвижимости нужна:
- Налоговой, чтобы подтвердить, что квартиры или дома у вас больше нет.
- Органу, который принимает решение о предоставлении жилья.
- Банку при решении вопроса о большом кредите.
Выписка о кадастровой стоимости требуется налоговой, чтобы проверить правильность начислений.
Отчет из ЕГРН онлайн официально из Росреестра за 15 минут
Отчет об объекте недвижимости на основе информации из ЕГРН
Отчет из ЕГРН о переходе прав на объект недвижимости
Заказать отчет
Полная информация об объекте недвижимости (2 отчета)
Заказать отчет
Отчет на квартиру, частный дом
Отчет на земельный участок
Отчет на здание, сооружение
Значение кадастровой выписки из ЕГРН?
Кадастровая отчет из ЕГРН – это справка выдаваемая по запросу любому лицу. Содержит данные об объекте недвижимости. Объектом выступают — квартира, дом или земля. Справка выдается на всю недвижимость, зарегистрированную на территории РФ. В ЕГРН заносятся данные об операциях с недвижимостью. Информация доступна и выдается всем желающим. Отчет из ЕГРН (ЕГРП) бесплатно не предоставляется, необходимо оплатить пошлину.
Содержит данные об объекте недвижимости. Объектом выступают — квартира, дом или земля. Справка выдается на всю недвижимость, зарегистрированную на территории РФ. В ЕГРН заносятся данные об операциях с недвижимостью. Информация доступна и выдается всем желающим. Отчет из ЕГРН (ЕГРП) бесплатно не предоставляется, необходимо оплатить пошлину.
Что я узнаю из выписки ЕГРН?
Информация, содержащаяся в отчете из ЕГРН зависит от вида документа.
› О переходе прав на объект недвижимости – сведения о собственниках, адресе объекта, форме права собственности, дате регистрации и прекращения прав, основании возникновения прав.
› О характеристиках и правах – данные о зарегистрированных правах, кадастровой стоимости, технические сведения. ЕГРН отчет содержит описание местоположения, план помещения на этаже и информацию об обременениях.
Официальная проверка недвижимости на основании онлайн сведений из Росреестра
ЕГРН ОНЛАЙН — коммерческий сервис, оказывающий помощь в получении официальных документов и сведений в Росреестре и кадастровой палате.
Сервис позволяет сформировать и направить запрос к информационному ресурсу Федеральной службы государственной регистрации, кадастра и картографии РФ и получить ответ в виде файла, который формируется в отчет и автоматически перенаправляется на указанный Вами адрес электронной почты. Также вам отправляется и отчет ЕГРН для подтверждения легитимности информации, отображаемой в отчете. В работе сервиса мы используем открытое API Федеральной службы государственной регистрации, кадастра и картографии РФ, а также руководствуемся рекомендациями Росреестра по работе с информационными ресурсами. Технологии использования данных ресурсов не подлежат лицензированию и свободно размещаются Росреестром в открытом доступе.
Согласно ст. 62 гл. 8 Федерального закона № 218-ФЗ «О государственной регистрации недвижимости», с 01.01.2017, получить информацию об объекте недвижимости в виде выписки из ЕГРН, может любой гражданин, без согласия собственника. Возможность получения электронной выписки через портал Росреестра регламентирована приказом Минэкономразвития от 23 декабря 2015 г. N968.
N968.
Стоит ли доверять отчете ЕГРН, которую предоставляет продавец?
В некоторых случаях продавец заинтересован в передаче ложной информации об объекте недвижимости. Например, при наличии обременения, ареста или в случае мошеннических действий. Ради продажи квартиры преступникам ничего не стоит подделать справку и выдать недвижимость за свою собственность. Рекомендуем самостоятельно заказать отчет ЕГРН Росреестра, чтобы быть уверенным в ее подлинности.
Нужно ли для совершения сделок свидетельство о праве собственности?
Свидетельство о регистрации права собственности оформлялось на недвижимость до 2017 года. В 2017 году документ стал не нужен, т.к. появилась отчет ЕГРН. Справка подтверждает право собственности и является официальным документом, который выдает Росреестр. Отчет ЕГРН из Росреестра онлайн оформляется за 15 минут на нашем сайте.
Стоит ли беспокоиться о легитимности электронной выписки из Росреестра?
Электронная отчет обладает такой же юридической силой, как и бумажная. Документы идентичны, содержат одинаковую информацию, заверяются подписями Росреестра. Электронную отчет ЕГРН выдает официальный сайт учреждения. Сомневаться в легитимности нет причин. Обращаем внимание, электронная подпись — это не синяя печать, это файл.
Документы идентичны, содержат одинаковую информацию, заверяются подписями Росреестра. Электронную отчет ЕГРН выдает официальный сайт учреждения. Сомневаться в легитимности нет причин. Обращаем внимание, электронная подпись — это не синяя печать, это файл.
Что такое Росреестр и кадастровая палата и чем они отличаются?
Росреестр — орган исполнительной власти. Кадастровая палата ─ подведомственное учреждение Росреестра. Орган регистрирует права на объекты недвижимости и сделки с ним.
Зачем нужна кадастровая карта Росреестра?
Публичная кадастровая карта – это онлайн карта, на которую нанесена разметка в соответствии с выделенными объектами недвижимости. Ресурс хранит информацию об участках, домах, поставленных на кадастровый учет в ЕГРН и для которых проведено межевание. Кликнув на любой участок карты, вы узнаете кадастровый номер, площадь объекта, назначение, год постройки дома. Вы можете заказать отчет из ЕГРН на любой участок. Сведения на карте регулярно обновляются и дополняются через сервис Росреестр онлайн.
Что такое ЕГРП?
Ранее существовало два учреждения. ЕГРП — Единый государственный реестр прав на недвижимое имущество и ГКН — Государственный кадастр недвижимости. В 2017 году кадастр и реестр объединены в ЕГРН.
Помощь и поддержка — ответы на часто задаваемые вопросы
Откуда Вы берёте отчеты?
Мы предоставляем отчеты, на основании выписок из официальной онлайн базы данных Росреестра.
Сколько времени готовится отчет?
Это зависит от технической загруженности сервисов Росреестра. По нашей статистике, в 70% — до 3 часов, в 10% — до 24 часов. В исключительных случаях — до 48 часов.
По закону (п. 9 ст. 62 закона № 218-ФЗ), сведения из ЕГРН предоставляются в срок не более 3 рабочих дней.
Зачем нужен отчет на основе выписки из ЕГРН?
Сегодня выписка ЕГРН это обязательный документ для совершения любых сделок с недвижимостью. Он необходим для подтверждения прав собственника, проверки отсутствий обременений. Это главный способ проверки недвижимости перед покупкой и способ обезопасить себя от мошенников.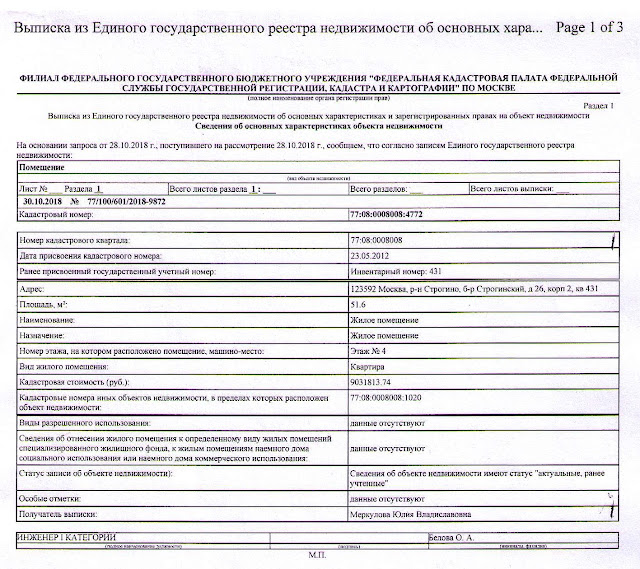 Выписка так же требуется как официальный документ для предоставления в суды, налоговые, для проведения кадастровых работ. Отчет позволит Вам ознакомиться оперативно с данными, которые есть в базе ЕРН Росреестра.
Выписка так же требуется как официальный документ для предоставления в суды, налоговые, для проведения кадастровых работ. Отчет позволит Вам ознакомиться оперативно с данными, которые есть в базе ЕРН Росреестра.
В каких случаях обязательна требуется выписка ЕГРН?
Вам обязательно потребуется выписка ЕГРН, если:
— Вступаете в наследство на недвижимость
— При регистрации сделки купли / продажи и подачи документов в Росреестр
— Оформлении ипотеки на квартиру, земельный участок
— Для судебных разбирательств
— Для уточнения границ земельного участка
— Бракоразводный процесс
— Деление имущества
— Встали в очередь на улучшение жилищных условий»
В чем плюсы отчета в электронном виде?
Отчет в электронном виде можно заказать в считанные минуты, не вставая из-за компьютера. Не нужно стоять в очередях в МФЦ, заполнять квитанции и заявления.
Как узнать кадастровый номер земельного участка?
Чтобы найти кадастровый номер земельного участка — нужно вбить в строку поиска его точный адрес и в результатах выдачи найти свой объект.
Другой вариант найти документы на землю, например паспорт объекта или свидетельство о регистрации собственности (зеленка).
Как узнать кадастровый номер квартиры, дома или сооружения?
Кадастровый номер содержится в документах на квартиру, кадастровом паспорте, свидетельстве. Так же вы можете найти кадастровый номер через форму поиска, для этого введите адрес квартиры и в результатах выдачи будет ваша квартира.
Если в поиске нет нужной квартиры или земельного участка?
Скорее всего вы ищете объект по адресу, если поиск не находит, то единственный вариант искать через кадастровый номер квартиры или участка. Если вам нужна выписка, но вы не можете найти объект на сайте, то напишите специалистам на почту [email protected] и вам помогут.
Почему в выписке ЕГРН отсутствуют данные о ФИО владельца?
Данные о владельце имеются только для объектов, которые были приобретены с 1998 года, так как до этого Росреестр не вёл единую базу.
Так же бывают исключения, например, для коммунальных квартир, нужно заказывать отдельную выписку на каждую комнату, так как у каждой комнаты разные владельцы.
Недавно оформленные права на недвижимость. Онлайн база ЕГРН обновляется не моментально, и недавно (2-3 недели) оформленные сделки там не отображаются. Связано это с тем, что региональные филиалы Росреестра передают информацию в Москву, а затем информация вносится в онлайн базу ЕГРН.
Иногда БТИ или Росреестр выделяет, к примеру, помещения в жилом частном доме, как самостоятельные объекты недвижимости и присваивает им кадастровые номера. У таких объектов также не будет записей о собственниках.
Если отчет не пришел на E-mail в течение 24 часов?
Проверьте папку «Спам» — иногда письма попадают туда, так как отправляются автоматически. Если ничего нет, то напишите ваш номер заявки в службу тех поддержки и Вам обязательно помогут найти выписку
Срок получения выписок зависит только от технической загруженности сервисов Росреестра, если Вы пока не получили выписку — это значит что Росреестр еще не обработал Ваш запрос. По закону, Росреестр обязан обработать запрос в срок не более 3 рабочих дней (п. 9 ст. 62 закона № 218-ФЗ), ожидайте, пожалуйста. Если выписка не пришла по окончанию этого срока, мы вернём Вам деньги.
9 ст. 62 закона № 218-ФЗ), ожидайте, пожалуйста. Если выписка не пришла по окончанию этого срока, мы вернём Вам деньги.
Обращаем Ваше внимание, что ВСЕ сервисы предлагающие услуги по получению выписок из ЕГРН работают с одной и той же базой — базой Росреестра и скорость получения выписки у всех зависит только от него.
Я заказывал(а) две выписки, а пришла только одна?
Два типа выписок приходят двумя письмами. Они могут приходить с некоторой разницей во времени, это зависит от технической загруженности сервисов Росреестра.
В выписке некорректные или устаревшие данные?
Через наш сервис Вы подаете запрос и получаете сведения имеющиеся в базе Росреестра на момент запроса. Обновление баз данных зависит от оперативности региональных подразделений Росреестра. По нашим наблюдениям, при регистрации нового объекта или изменений в существующих, срок обновления данных составляет в среднем от одной до двух недель. Если Вы находите полученную информацию некорректной — обратитесь в региональное подразделение Росреестра или по телефону горячей линии 8 (800) 100-34-34.
У меня другая проблема!
Если ничего из вышеперечисленного не прояснило ситуацию, напишите нам.
Внимание!
Перед тем как писать, убедитесь, что выписка была заказана именно у нас, на rosegrp.ru. Мы не отвечаем за выписки, заказанные на других сайтах. В письме обязательно укажите номер транзакции, дату заказа выписки и E-mail, на который она была заказана. Обращения без этих данных игнорируются. Электронная почта технической поддержки сервиса:
[email protected].
Росреестр рассказал, в каких случаях нужны выписки из ЕГРН
Алексей Филиппов/ РИА Новости
Выписка необходима при проведении сделок с недвижимостью (например, чтобы подтвердить законность владения объектом), при использовании объекта в качестве залога, оспаривании сделок в суде, открытии наследства, оформлении завещания и др. При этом бывают разные виды выписок. Они подразделяются на те, которые предоставляются по запросу любого заинтересованного лица, и те, которые доступны только ограниченному кругу лиц, установленному законом (собственникам или их законным представителям, правоохранительным органам, судам, судебным приставам, нотариусам, органам власти).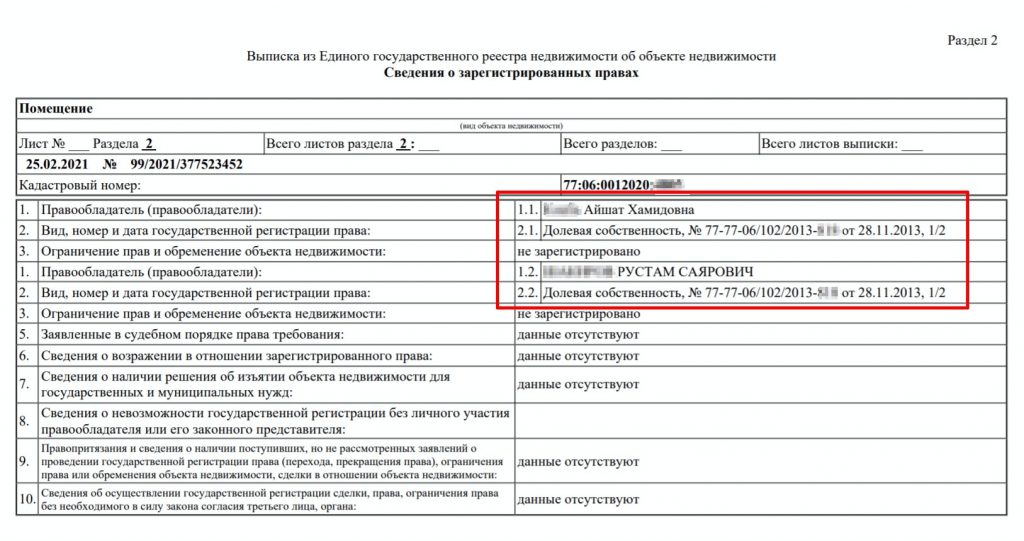 Это зависит от того, какие сведения в них содержатся — общедоступные или ограниченного доступа.
Это зависит от того, какие сведения в них содержатся — общедоступные или ограниченного доступа.
К общедоступным сведениям относятся сведения о характеристиках объекта, зарегистрированных правах на него, о переходе прав на объект (при этом в отношении правообладателя в этих случаях указываются неполные сведения — например, только фамилия, имя, отчество, если собственником является физическое лицо). К сведениям закрытого доступа относятся сведения о правах отдельного лица на имеющиеся у него объекты недвижимости, о содержании правоустанавливающего документа, о признании правообладателя недееспособным или ограниченно дееспособным, о лицах, получивших сведения об объекте недвижимости. Также ограничено предоставление копий документов, на основании которых в ЕГРН были внесены сведения о недвижимости (например, копия межевого, технического плана).
Выписка об основных характеристиках и зарегистрированных правах на объект недвижимости может понадобиться тому, кто хочет подтвердить свои права на недвижимость, а также тем, кому необходимо получить информацию о наличии зарегистрированных обременений на объект. В ней содержится информация о собственнике (при этом если заявителем является иное лицо, не собственник или его представитель, то выписка не будет содержать полных сведений о собственнике, а будут указаны только, например, фамилия, имя и отчество), адресе объекта, его кадастровой стоимости, дате ввода объекта в эксплуатацию (завершения строительства).
Здесь же можно посмотреть сведения о возможном ограничении прав и обременении объекта (например, если на квартиру наложен арест по решению суда или объект приобретен в ипотеку), план расположения помещения на этаже, схематическое отображение расположения объекта на земельном участке. Также в данной выписке можно проверить наличие отметки о согласии или отсутствии согласия супруга на продажу объекта недвижимости.
Выписка из ЕГРН об объекте недвижимости содержит наиболее полную информацию о характеристиках объекта недвижимости. Например, она поможет определить, попадает ли земельный участок в границы охранной зоны или зоны с особыми условиями использования территории, а также узнать, включен ли объект в реестр объектов культурного наследия. В нее включено описание местоположения границ объекта, сведения об ограничениях его использования, а также о наличии или отсутствии ранее возникших прав (то есть прав, которые возникли до 31 января 1998 года).
Выписка из ЕГРН о переходе прав на объект понадобится, чтобы узнать полную историю владения объектом. В ней есть информация как о текущем владельце, так и о предыдущих собственниках. Указываются даты регистрации переходов права собственности и вид документа, на основании которого был зарегистрирован такой переход права.
Выписка из ЕГРН о зарегистрированных договорах участия в долевом строительстве нужна тем, кто приобретает квартиру или нежилое помещение по уступке права требований. Потенциальный участник долевого строительства с ее помощью сможет выяснить, сколько объектов уже продано в конкретном строящемся доме. Документ представляет собой выписку о земельном участке, на котором ведется строительство многоквартирного дома. Застройщик в ней указывается как правообладатель земельного участка, а зарегистрированные ДДУ перечисляются в качестве обременений на сам участок.
Выписка из ЕГРН о кадастровой стоимости объекта недвижимости поможет собственникам узнать сведения о кадастровой стоимости объекта и проверить правильность расчета налога на имущество. Документ содержит расширенную информацию о величине кадастровой стоимости на дату, указанную в запросе; об акте, на основании которого определена кадастровая стоимость; о кадастровом номере объекта, о датах утверждения и применения кадастровой стоимости.
Есть также целый ряд выписок со сведениями ограниченного доступа. Это, в частности, выписка из ЕГРН о правах отдельного лица на имевшиеся или имеющиеся у него объекты недвижимости. Такая выписка может пригодиться в случае, если вам продолжает приходить уведомление об уплате налога за недвижимость, которая была продана.
В выписке содержатся сведения о наличии прав на недвижимость по состоянию на определенную дату, если правообладатель ее указал. Документ позволяет подтвердить, какой недвижимостью владел правообладатель в течение конкретного периода. Выписку из ЕГРН о признании правообладателя недееспособным или ограниченно дееспособным нередко необходимо предоставлять в суд или нотариусу в качестве подтверждения, что участник сделки является дееспособным. Это обусловлено тем, что сделка, совершенная с недееспособным правообладателем, будет считаться недействительной.
Справка о лицах, получивших сведения об объекте недвижимости, может понадобиться, если правообладателю необходима информация о том, кто интересовался его собственностью. В справке будут указаны физические и юридические лица или органы местного самоуправления, которые получали сведения об объекте, дату получения ими сведений и исходящий номер выписки.
Наконец, выписка о содержании правоустанавливающих документов пригодится, если, например, оригиналы документов на жилье утрачены. В ней можно получить информацию о содержании документа, на основании которого возникло право собственности.
Получить выписку из ЕГРН можно на объект в любом регионе России, ехать туда не обязательно. Бумажный вариант выписки можно получить через МФЦ. Выписку в электронной форме можно заказать на сайте Росреестра или Федеральной кадастровой палаты. С мая 2021 года получить наиболее востребованные виды выписок из ЕГРН также можно на портале госуслуг. Среди них выписка об основных характеристиках и зарегистрированных правах на объект недвижимости; об объекте недвижимости; о переходе прав на объект недвижимости.
В интернете немало сайтов-двойников Росреестра, которые предлагают услуги предоставления выписок из ЕГРН от лица Росреестра и имеют схожие с официальным сайтом символику и название, отмечают в ведомстве. Заказывать выписки нужно только на официальных сайтах Росреестра, Федеральной кадастровой палаты, на портале госуслуг или обратившись в МФЦ.
Сведения ЕГРН предоставляются за плату. Размер платы зависит от вида запрашиваемой информации, формы предоставления сведений (на бумажном носителе или в виде электронного документа), статуса заявителя (физическое или юридическое лицо). Бесплатно эти сведения представляются по запросам правоохранительных органов, судов, судебных приставов, прокуратуры, органов власти, Банка России, госкорпораций «Росатом» и «Роскосмос».
Бесплатно эти сведения представляются по запросам правоохранительных органов, судов, судебных приставов, прокуратуры, органов власти, Банка России, госкорпораций «Росатом» и «Роскосмос».
Также бесплатное предоставление сведений ЕГРН предусмотрено по запросам председателя Счетной палаты РФ и контрольно-счетных органов в субъектах, арбитражных управляющих, Уполномоченного при Президенте РФ по защите прав предпринимателей, Уполномоченного при Президенте РФ по правам ребенка, Пенсионного фонда РФ, Центральной избирательной комиссии РФ и избирательных комиссий в субъектах и муниципалитетах, если эти сведения необходимы для осуществления их полномочий.
Кроме того, на бесплатной основе сведения из реестра недвижимости могут получать многофункциональные центры для предоставления государственных или муниципальных услуг и нотариусы.
Чем различаются основные виды выписок из ЕГРН.
-
Главная - /
-
Новости - /
-
Общие новости
Выписки из Единого государственного реестра недвижимости (ЕГРН) используются для подтверждения права собственности при проведении сделок, определении налоговых обязательств собственников, использовании объекта в качестве залога, оспаривании сделок в суде, открытии наследства, оформлении завещания и т. д.
д.
Выписки из ЕГРН отличаются друг от друга содержанием, формой, стоимостью и даже возможностью их получения. Для начала рассмотрим несколько основных видов выписок сведений из ЕГРН, относящихся к общедоступным.
- Выписка об основных характеристиках и зарегистрированных правах на объект недвижимости
Этот вид выписки официально подтверждает, что в ЕГРН содержатся сведения о кадастровом учете интересующего объекта недвижимости и зарегистрированных на него правах. Данная выписка содержит такие данные, как кадастровый номер объекта и дата его присвоения, адрес, площадь, назначение; информацию о правообладателях, видах права, номере и дате регистрации, наличии ограничений прав или обременений. Такая выписка содержит описание местоположения объектов и план расположения помещений, машино-мест в здании, данные о кадастровой стоимости, характерных точках границ и т.п.
- Выписка об объекте недвижимости
Выписка является расширенным вариантом выписки об основных характеристиках и зарегистрированных правах на объект недвижимости и содержит наиболее полную информацию об объекте. Кроме общей информации, данный вид выписки содержит обязательное описание местоположения границ объекта, сведения об ограничениях его использования, а также о правах, возникших на него, но не зарегистрированных до 1998 года. Только в этой выписке можно получить сведения о том, попадает ли земельный участок в границы охранной зоны или зоны с особыми условиями использования территории, а также узнать, включена ли недвижимость в реестр объектов культурного наследия.
- Выписка о переходе прав
Выписка содержит информацию не только о текущем владельце, но и о предыдущих – с указанием дат регистрации предыдущих переходов права и документов-оснований. Стоит внимательно отнестись к интересующему объекту недвижимости, если в отношении этого объекта часто совершались сделки и сменялись собственники. Это может косвенно свидетельствовать о скрытых проблемах, связанных с конкретным объектом недвижимости.
- Выписка о кадастровой стоимости объекта недвижимости
Данный документ имеет четкое целевое назначение и позволяет получить актуальные данные о кадастровой стоимости объекта. Применение указанной справки может заключаться в определении рыночной цены объекта, проверке правильности расчета налога на имущество.
Помимо общедоступной информации в ЕГРН хранится информация, получить которую могут только собственники или их законные представители, а также правоохранительные органы, суды, приставы, нотариусы, органы власти. К сведениям ограниченного доступа относятся следующие основные виды выписок.
- Выписка о признании правообладателя недееспособным или ограниченно дееспособным
Справка с таким содержанием потребуется для проверки продавца недвижимости, чтобы удостовериться в дееспособности человека, ведь сделка, совершенная с недееспособным правообладателем будет считаться недействительной.
- Выписка о правах отдельного лица на имевшиеся (имеющиеся) у него объекты недвижимости
С помощью выписки можно получить сведения о наличии прав на недвижимость по состоянию на определенную дату.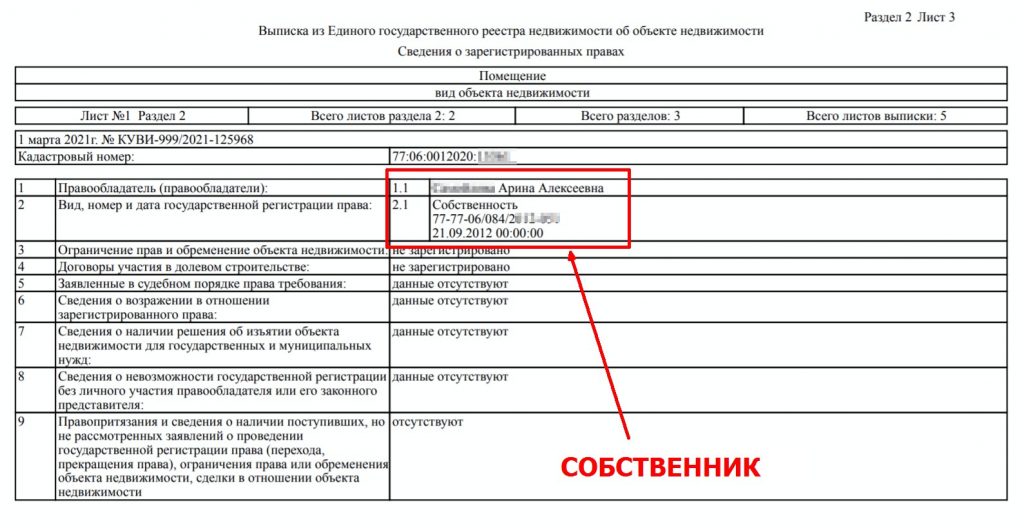 Позволяет подтвердить, какая недвижимость была зарегистрирована на вас в интересуемый период, в том числе за прошедшее время. Выписка нужна в ситуации, когда, например, после продажи квартиры вы получаете уведомление об уплате налога. Также выписка пригодится для оформления наследства, чтобы узнать о наличии недвижимости у наследодателя.
Позволяет подтвердить, какая недвижимость была зарегистрирована на вас в интересуемый период, в том числе за прошедшее время. Выписка нужна в ситуации, когда, например, после продажи квартиры вы получаете уведомление об уплате налога. Также выписка пригодится для оформления наследства, чтобы узнать о наличии недвижимости у наследодателя.
- Выписка о лицах, получивших сведения об объекте недвижимого имущества
Выписка предоставляет информацию о том, кто интересовался недвижимостью, которая находится у вас в собственности. В документе будут содержаться ФИО лиц, которые запрашивали сведения из ЕГРН в отношении вашего объекта недвижимости. Такая выписка позволяет выявить возможные мошеннические действия в отношении вашей недвижимости.
- Выписка о содержании правоустанавливающих документов
Правообладатели или законные представители запрашивают данную выписку, как правило, для предоставления в кредитные и страховые организации при совершении различного рода сделок с привлечением заемных средств. Пригодится, если оригиналы бумаг на жилье утрачены. Выписка сможет доказать наличие вашего права собственности и дать информацию о том, откуда право появилось и на каком основании сохраняется. Выписку часто запрашивают нотариусы для совершения нотариальных действий, связанных с открытием наследства, органы государственной власти и местного самоуправления при оказании государственных и муниципальных услуг, суды.
Пригодится, если оригиналы бумаг на жилье утрачены. Выписка сможет доказать наличие вашего права собственности и дать информацию о том, откуда право появилось и на каком основании сохраняется. Выписку часто запрашивают нотариусы для совершения нотариальных действий, связанных с открытием наследства, органы государственной власти и местного самоуправления при оказании государственных и муниципальных услуг, суды.
Как получить выписку из ЕГРН?
Получить выписку из ЕГРН можно несколькими способами: в офисах МФЦ, на официальном сайте Росреестра или посредством онлайн-сервиса Федеральной кадастровой палаты.
Сведения ЕГРН предоставляются в срок не более трех рабочих дней со дня получения органом регистрации прав запроса о предоставлении сведений. В случае подачи запроса через МФЦ срок предоставления сведений увеличивается на два дня.
Стоимость выписки варьируется в зависимости от вида предоставляемых сведений, формы документа и типа заявителя.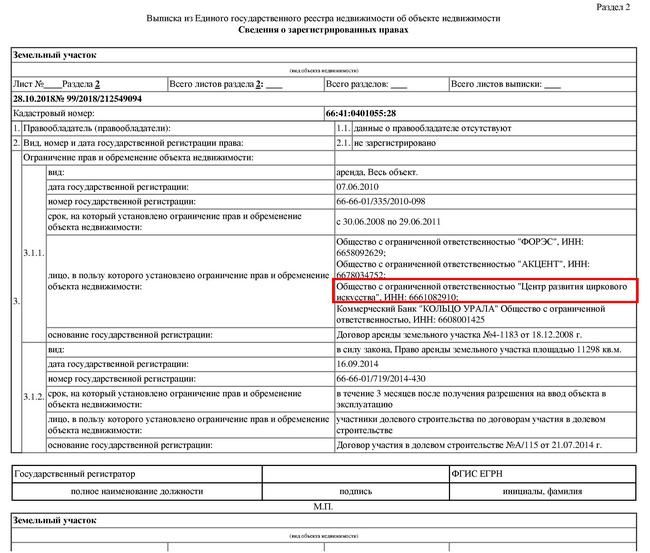 Выписка о кадастровой стоимости объекта недвижимости предоставляется бесплатно.
Выписка о кадастровой стоимости объекта недвижимости предоставляется бесплатно.
Создана: 11.02.2021 14:53, обновление 11.02.2021 14:53
Росреестр Онлайн – Выписка из ЕГРН (ЕГРП) за 5 минут
Что такое
выписка из ЕГРН?
Выписка из ЕГРН — это официальная справка с информацией о квартире, доме или земельном участке. С помощью этой справки любой желающий может узнать сведения о любой недвижимости,
находящейся на территории России, независимо от прав собственности на этот объект.
ЕГРН — Единый Государственный Реестр Недвижимости. В реестре хранятся все данные о квартирах, земельных участках и домах. Как только происходит какая-то операция с недвижимостью,
данные в обязательном порядке заносятся в реестр и становятся доступными для всех желающих. Таким образом государство защищает граждан от мошенничества при сделках с недвижимостью.
Для чего нужна
выписка ЕГРН?
Перед покупкой или продажей недвижимости важно убедиться, что площадь не находится в залоге, не арестована и не принадлежит постороннему лицу. Единственный законный способ это проверить —
Единственный законный способ это проверить —
заказать справку из ЕГРН.
Справка из ЕГРН также потребуется для оформления кредита, наследства или налогового вычета на квартиру, во время приватизации или судебного разбирательства, а также для других операций
с недвижимостью.
Что я узнаю из
выписки ЕГРН?
В выписке из ЕГРН вы найдете основные технические данные и сведения о зарегистрированных правах на квартиру, дом или участок:
кадастровую стоимость, описание местоположения, план помещения на этаже, действующих и прошлых владельцах, форму права и обременения.
Какие бывают
выписки из ЕГРН?
Для проверки квартиры, земельного участка, дома и другой недвижимости существует две основные справки:
- Справка из ЕГРН об объекте недвижимости
- Справка из ЕГРН о переходе прав на недвижимость
Из справки об объекте недвижимости вы узнаете технические сведения, собственников, а также наложены ли на квартиру, дом или участок обременения,
аресты и другие ограничения.
Из справки о переходе прав на недвижимость вы узнаете полную историю всех собственников. Эта информация поможет как минимум в двух случаях:
- Если квартира часто переходила от одного собственника к другому несколько раз — что-то здесь не так. Проверьте собственников.
- На права на площадь может претендовать другой человек по наследству (о чем вы можете не знать). Если на квартиру, дом или участок претендует
наследник, то оспорить свое право на жилье он может только в течение трех лет. Проверьте, сколько времени недвижимость принадлежала каждому собственнику.
Какой срок действия у
выписки из ЕГРН?
Срок справки из ЕГРН законом не ограничен, но банки, нотариусы и налоговая, обычно, работают с документами не старше одного месяца. Чем справка новее, тем больше
к ней доверия, ведь с момента её оформления сведения о недвижимости могли измениться.
Например, 1 сентября вы сделали выписку на квартиру — в ней указан владелец Иванов. 7 сентября вы решаете купить эту квартиру, но за этот момент Иванов
уже продал квартиру другому собственнику, сведения в Росреестре обновились. В такой ситуации ваша справка будет иметь устаревшие данные, о которых вы узнаете только
когда получите новую выписку.
Можно ли доверять чужим справкам?
Мы не рекомендуем доверять справкам, сделанным действующим собственником недвижимости (или тем, кто теоретически может быть заинтересован в обмане). Всегда заказывайте выписку самостоятельно. Только в этом
случае вы можете быть уверены в достоверности информации.
Заменяет ли
выписка из ЕГРН свидетельство права собственности?
Да, заменяет. Свидетельство о регистрации права собственности выдавали только до 2016 года. После этот документ заменили выпиской из ЕГРН. Выписка из ЕГРН —
Выписка из ЕГРН —
официальный документ Росреестра, подтверждающий права на собственность и заявленные при сделке данные.
Чем электронная
выписка из ЕГРН отличается от бумажной?
Бумажная и электронная версии документа ничем не отличаются юридически. Бумажный документ заверен ручной подписью, электронный — цифровой подписью Росреестра (файл с расширением
.sig). Оба метода заверки документа одинаково законны.
Разница только в удобстве. За бумажной выпиской придется ехать в офис Росреестра, стоять в очереди и ждать справку неделю. Электронную же выписку можно заказать
на нашем сайте не выходя из дома. При этом выписка будет оформлена за полчаса.
Могу ли я получить
выписку из ЕГРН на квартиру в другом городе?
Да, можете. Ваше местонахождение, прописка и право собственности на интересующий объект недвижимости никак не влияют на возможность заказать выписку. Выписку из ЕГРН может
Выписку из ЕГРН может
получить любой гражданин на любую недвижимость на территории России.
Белки ответа раннего роста (EGR) и ядерные факторы активированных Т-клеток (NFAT) образуют гетеродимеры и регулируют экспрессию генов провоспалительных цитокинов
1. Müller H.-J., Skerka,C., Bialonski,A. и Ципфель, П.Ф. (1991) Клон pAT 133 идентифицирует ген, который кодирует другой человеческий член класса генов, индуцированных фактором роста, с почти идентичными доменами цинковых пальцев. проц. Натл акад. науч. США, 88, 10079–10083. [Бесплатная статья PMC] [PubMed] [Google Scholar]
2. Perez-Castillo A., Pipaon,C., Garcia,I. и Алемани, С. (1993) Экспрессия гена NGFI-A необходима для пролиферации Т-лимфоцитов. Дж. Биол. Chem., 268, 19445–19450. [PubMed] [Google Scholar]
3. Гашлер А., Сухатме В.П. (1995) Белок 1 ответа на ранний рост (Egr-1): прототип семейства факторов транскрипции с цинковыми пальцами. прог. Нуклеиновая Кислота Рез. Мол. биол. , 50, 191–224. [PubMed] [Google Scholar]
, 50, 191–224. [PubMed] [Google Scholar]
4. Бекманн М.А. и Уилс П.А. (1997) Факторы транскрипции Egr в нервной системе. Нейрохим. Междунар., 31, 477–510. [PubMed] [Академия Google]
5. Milbrandt J. (1987) Ген, индуцированный фактором роста нервов, кодирует возможный фактор регуляции транскрипции. Наука, 238, 797–799. [PubMed] [Google Scholar]
6. Suggs S.V., Katzowitz, JL, Tsai-Morris, C. и Сухактме В.П. (1990) Последовательность кДНК гена реакции раннего роста клеток человека Egr-1. Nucleic Acids Res., 18, 4283. [Бесплатная статья PMC] [PubMed] [Google Scholar]
7. Райт Дж.Дж., Гюнтер К.С., Мицуя Х., Ирвинг С.Г., Келли К. и Зибенлист, У. (1990) Экспрессия гена цинкового пальца в клетках, трансформированных HTLV-I и HTLV-II. Наука, 248, 588–591. [PubMed] [Google Scholar]
8. Chavrier P., Zerial, M., Lemaire, P., Almendral, J., Bravo, R. и Чарни, П. (1988) Ген, кодирующий белок с цинковыми пальцами, активируется во время перехода G0/G1 в культивируемых клетках. EMBO J., 7, 29–35. [Бесплатная статья PMC] [PubMed] [Google Scholar]
EMBO J., 7, 29–35. [Бесплатная статья PMC] [PubMed] [Google Scholar]
и Сухатме В.П. (1988) Молекулярное колонирование, секвенирование и картирование EGR2, гена реакции раннего роста человека, кодирующего белок со структурой «цинк-связывающего пальца». проц. Натл акад. науч. США, 85, 7164–7168. [Бесплатная статья PMC] [PubMed] [Google Scholar]
10. Маги Х.В., Штаммингер Т., Рильке О., Браво Р. и Крочек Р.А. (1993) Экспрессия Pilot, предполагаемого фактора транскрипции, требует двух сигналов и чувствительна к циклоспорину А в Т-клетках. Междунар. Иммунол., 5, 63–70. [PubMed] [Google Scholar]
11. Патвардхан С., Гашлер А., Сигель М.Г., Чанг Л.К., Джозеф Л., Шоу Т.Б., Ле Бо М.М. и Сухатме В.П. (1991) EGR-3, новый член семейства генов Egr, кодирующих факторы немедленной ранней транскрипции. Онкоген, 6, 917–928. [PubMed] [Google Scholar]
12. Кросби С.Д., Путц Дж.Дж., Симбургер К.С., Фарнер Т.Дж. и Милбрандт, Дж. (1991) Ген раннего ответа NGFI-C кодирует активатор транскрипции цинковых пальцев и является членом семейства белков, связывающих элемент GCGGGGGCG (GSG). Мол. Клетка. биол., 11, 3835–3841. [Бесплатная статья PMC] [PubMed] [Google Scholar]
13. Cao X., Guy,G.R., Sukhatme,V.P. и Тан, Ю.Х. (1993) Регуляция гена EGR-1 фактором некроза опухоли и интерферонами в первичных фибробластах человека. Дж. Биол. Chem., 267, 1345–1349. [PubMed] [Google Scholar]
14. Скерка С., Декер, Э.Л. и Ципфель, П.Ф. (1995) Регуляторный элемент в промоторе гена интерлейкина 2 человека представляет собой сайт связывания белков цинковых пальцев Sp1 и EGR-1. Дж. Биол. хим., 270, 22500–22506. [PubMed] [Google Scholar]
15. Кремер Б., Мейхле А., Хенсел Г., Чарней П. и Крёнке, М. (1994) Характеристика чувствительного элемента Krox-24/Egr-1 в промоторе фактора некроза опухоли человека. Биохим. Биофиз. Акта, 1219 г., 413–421. [PubMed] [Google Scholar]
16. Лин Дж.-Х. и Леонард, У.Дж. (1997) Продукт немедленного раннего гена Egr-1 регулирует промотор b-цепи рецептора интерлейкина-2 человека через неканонические сайты связывания Egr и Sp1. Мол. Клетка. биол., 17, 3714–3722. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Клетка. биол., 17, 3714–3722. [Бесплатная статья PMC] [PubMed] [Google Scholar]
17. Dinkel A., Aicher, W.K., Haas, C., Zipfel, P.F., Peter, H.H. и Эйбель, Х. (1997) Активность фактора транскрипции Egr-1 подавляет экспрессию Fas и CD23 в В-клетках. Дж. Иммунол., 159, 2678–2684. [PubMed] [Google Scholar]
18. Мальцман Дж.С., Кармен Дж.А. и Монро, Дж.Г. (1996) Регуляция транскрипции гена Icam-1 в В-лимфоцитах, стимулированных антигенным рецептором и форболовым эфиром: роль фактора транскрипции EGR-1. Дж. Эксп. Мед., 183, 1747–1759. [Бесплатная статья PMC] [PubMed] [Google Scholar]
19. Li-Weber M., Laur, O. и Краммер, П.Х. (1999) Новые композитные сайты Egr/NFAT опосредуют активацию промотора лиганда CD95 (APO/Fas) в ответ на стимуляцию Т-клеток. Евро. Дж. Иммунол., 29 лет, 3017–3027. [PubMed] [Google Scholar]
20. Кристи Б. и Натанс Д. (1989) Сайт связывания ДНК белка Zif268, индуцируемого фактором роста. проц. Натл акад. науч. США, 86, 8737–8745. [Бесплатная статья PMC] [PubMed] [Google Scholar]
21. Ван З.Ю. и Дуэль, Т.Ф. (1992) Чувствительный к нуклеазе S1 гомопуриновый/гомопиримидиновый домен в промоторе А-цепи PDGF содержит новый сайт связывания для индуцируемого фактором роста белка EGR-1. Биохим. Биофиз. Рез. Комм., 188, 433–439. [PubMed] [Академия Google]
Ван З.Ю. и Дуэль, Т.Ф. (1992) Чувствительный к нуклеазе S1 гомопуриновый/гомопиримидиновый домен в промоторе А-цепи PDGF содержит новый сайт связывания для индуцируемого фактором роста белка EGR-1. Биохим. Биофиз. Рез. Комм., 188, 433–439. [PubMed] [Академия Google]
22. Зипфель П.Ф., Декер Э.Л., Холст С. и Скерка, С. (1997) Человеческий белок цинковых пальцев EGR-4 действует как ауторегуляторный репрессор транскрипции. Биохим. Биофиз. Акта, 1354, 134–144. [PubMed] [Google Scholar]
23. Бесиада Э., Разанди М. и Левин, Э.Р. (1996) Egr-1 активирует транскрипцию основного фактора роста фибробластов. Дж. Биол. Chem., 271, 18576–18581. [PubMed] [Google Scholar]
24. Ху Р.М. и Левин, Э.Р. (1994) Рост астроцитов регулируется нейропептидами через Tis 8 и основной фактор роста фибробластов. Дж. Клин. Инвест., 193, 1820–1827 гг. [Бесплатная статья PMC] [PubMed] [Google Scholar]
25. Хачигян Л.М., Линднер В., Уильямс А.Дж. и Коллинз, Т. (1996) Egr-1-индуцированная экспрессия эндотелиальных генов: общая тема при повреждении сосудов. Наука, 271, 1427–1430. [PubMed] [Google Scholar]
Наука, 271, 1427–1430. [PubMed] [Google Scholar]
26. Лю Дж., Гроган Л., Нау М.М., Аллегра С.Дж., Чу Э. и Райт, Дж.Дж. (2001)Физическое взаимодействие между p53 и геном первичного ответа Egr-1. Междунар. J. Oncol., 18, 863–870. [PubMed] [Google Scholar]
27. Cui M.-Z., Party, GCN, Oeth, P., Larson, H., Smith, M., Huang, R.-P., Adamson, E.D. и Макман, Н. (1996) Транскрипционная регуляция гена тканевого фактора в эпителиальных клетках человека опосредована Sp1 и EGR-1. Дж. Биол. Chem., 271, 2731–2739. [PubMed] [Google Scholar]
28. Lui C., Adamson,E. и Меркола, Д. (1996) Фактор транскрипции EGR-1 подавляет рост и трансформацию клеток фибросаркомы HAT-1080 человека путем индукции трансформирующего фактора роста b1. проц. Натл акад. науч. США, 93, 11831–11836. [Бесплатная статья PMC] [PubMed] [Google Scholar]
29. Day ML, Wu, S. и Баслер, Дж.В. (1993) Ген, индуцируемый фактором роста простатического нерва А, связывает новый элемент в промоторе гена ретинобластомы. Рак Res., 53, 5597–5599. [PubMed] [Google Scholar]
Рак Res., 53, 5597–5599. [PubMed] [Google Scholar]
30. Филипп А., Шнайдер А., Васрик И., Финке К., Сюн Ю., Беш Д., Алитало К. и Эйлерс, Э. (1994)Репрессия циклина D1: новая функция MYC. Мол. Клетка. биол., 14, 4032–4043. [Бесплатная статья PMC] [PubMed] [Google Scholar]
31. Tremblay J.J. и Друэн, Дж. (1999) Egr-1 является нижестоящим эффектором GnRH и синергизируется за счет прямого взаимодействия с Ptx1 и SF-1 для усиления транскрипции гена лютеинизирующего гормона β. Мол. Клетка. биол., 19, 2567–2576. [PMC free article] [PubMed] [Google Scholar]
32. Ли С.Л., Садовский Ю., Свирнов Х., Полиш Ю.А., Года П., Гаврилина Г. и Милбрандт, Дж. (1996) Дефицит лютеинизирующего гормона и бесплодие самок у мышей, лишенных фактора транскрипции NGFI-A (Egr-1). Наука, 273, 1219–1221. [PubMed] [Google Scholar]
33. Шнайдер-Монури С., Топилка П., Сейтанду Т., Леви Г., Коэн-Таннуджи М., Пурнин С., Бабине С. и Чарней, П. (1993) Разрушение Krox-20 приводит к изменению ромбомеров 3 и 5 в развивающемся заднем мозге. Сотовая, 75, 1199–1214. [PubMed] [Google Scholar]
Сотовая, 75, 1199–1214. [PubMed] [Google Scholar]
34. Tourtellotte WG and Milbrandt, J. (1998) Сенсорная атаксия и агенезия мышечного веретена у мышей, лишенных транскрипционного фактора Egr3. Nature Genet., 20, 87–91. [PubMed] [Google Scholar]
35. Tourtellotte W.G., Nagarajan, R., Auyeung, A., Mueller, C. и Милбрандт, Дж. (1999) Бесплодие, связанное с неполной остановкой сперматогенеза и олигоспермией у мышей с дефицитом Egr-4. Развитие, 126, 5061–5071. [PubMed] [Google Scholar]
36. Cogswell PC, Mayo, M.W. и Болдуин, А.С., младший (1997) Участие синергизма Egr-1/RelA в различении активации Т-клеток от транскрипции NF-kB1, индуцированной фактором некроза опухоли-α. Дж. Эксп. Мед., 185, 491–497. [Бесплатная статья PMC] [PubMed] [Google Scholar]
37. Silverman E.S., Du, J., Williams, A.J., Wadgaonkar, R., Drazen, J.M. и Collins T. (1998) цАМФ-ответный элемент-связывающий белок-связывающий белок (CBP) и p300 являются коактиваторами транскрипции раннего фактора роста-1 (Egr-1). Биохим. Дж., 336, 183–189. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Биохим. Дж., 336, 183–189. [Бесплатная статья PMC] [PubMed] [Google Scholar]
38. Сривастава С., Вайцман М.Н., Кимбл Р.Б., Риццо М., Занер М., Милбрандт Дж., Росс Ф.П. и Пасифичи, Р. (1998) Эстроген блокирует экспрессию гена M-CSF и образование остеокластов, регулируя фосфорилирование Egr-1 и его взаимодействие с Sp1. Дж. Клин. Инвест., 102, 1850–1859. [Бесплатная статья PMC] [PubMed] [Google Scholar]
39. Yang Y., Dong, B., Mittelstadt, P.R., Xiao, H. и Эшвелл, Дж.Д. (2002) HIV Tat связывает белки Egr и усиливает Egr-зависимую трансактивацию промотора лиганда Fas. Дж. Биол. хим., 277, 19482–19487. [PubMed] [Google Scholar]
40. Trejo S.R., Fahl,W.E. и Ратнер, Л. (1997) Белок Tax вируса Т-клеточного лейкоза человека типа 1 опосредует трансактивацию промотора c-sis/тромбоцитарного фактора роста-B посредством взаимодействия с факторами транскрипции цинковых пальцев Sp1 и NGFI-A/Egr-1. Дж. Биол. Chem., 272, 27411–27421. [PubMed] [Google Scholar]
41. Yoo Y. D., Ueda, H., Park, K., Flanders, K.C., Lee, Y.I., Jay, G. и Ким, С.-Дж. (1996) Регуляция экспрессии трансформирующего фактора роста-β1 трансактиватором X вируса гепатита В (HBV). Дж. Клин. Инвест., 97, 388–395. [Бесплатная статья PMC] [PubMed] [Google Scholar]
D., Ueda, H., Park, K., Flanders, K.C., Lee, Y.I., Jay, G. и Ким, С.-Дж. (1996) Регуляция экспрессии трансформирующего фактора роста-β1 трансактиватором X вируса гепатита В (HBV). Дж. Клин. Инвест., 97, 388–395. [Бесплатная статья PMC] [PubMed] [Google Scholar]
и Ким, С.-Дж. (1996) Регуляторный белок IE2 цитомегаловируса человека индуцирует экспрессию гена человеческого трансформирующего фактора роста β1 через сайт связывания Egr-1. J. Virol., 70, 7062–7070. [Бесплатная статья PMC] [PubMed] [Google Scholar]
43. Russo M., Sevetson, B.R. и Милбрандт, Дж. (1995) Идентификация Nab1, репрессора транскрипции, опосредованной NGFI-A и Krox20. проц. Натл акад. науч. США, 92, 6873–6877. [Бесплатная статья PMC] [PubMed] [Google Scholar]
и Милбрандт, Дж. (1996) NAB2, корепрессор NGFI-A (Egr-1) и Krox20, индуцируется пролиферативными и дифференциальными стимулами. Мол. Клетка. биол., 16, 3545–3553. [Бесплатная статья PMC] [PubMed] [Google Scholar]
45. Локсли Р.М., Киллин, Н. и Ленардо, М.Дж. (2001) Надсемейства TNF и рецепторов TNF: интеграция биологии млекопитающих. Ячейка, 104, 487–501. [PubMed] [Академия Google]
и Ленардо, М.Дж. (2001) Надсемейства TNF и рецепторов TNF: интеграция биологии млекопитающих. Ячейка, 104, 487–501. [PubMed] [Академия Google]
46. Декер Э.Л., Скерка С. и Ципфель, П.Ф. (1998) Белок ответа на ранний рост (EGR-1) регулирует транскрипцию интерлейкина-2 за счет синергетического взаимодействия с ядерным фактором активированных Т-клеток. Дж. Биол. Chem., 273, 26923–26930. [PubMed] [Google Scholar]
47. Рао А., Луо, К. и Хоган, П.Г. (1997) Факторы транскрипции семейства NFAT: регуляция и функция. Анну. Rev. Immunol., 15, 707–747. [PubMed] [Google Scholar]
48. Куклина Е.М., Ширшев С.В. (2001)Роль фактора транскрипции NFAT в иммунном ответе. Биохимия, 66, 467–475. [PubMed] [Академия Google]
49. Лопес-Родригес К., Арамбуру Дж., Ракеман А.С. и Рао, А. (1999) NFAT5, конститутивно ядерный белок NFAT, который не взаимодействует с Fos and Jun. Proc. Натл акад. науч. США, 96, 7214–7219. [Бесплатная статья PMC] [PubMed] [Google Scholar]
50. Skerka C. , Decker, E.L. и Ципфель, П.Ф. (1997) Координация экспрессии и различных характеристик ДНК-связывания четырех белков EGR-цинковых пальцев в Т-лимфоцитах Jurkat. Иммунобиология, 198, 179–191. [PubMed] [Академия Google]
, Decker, E.L. и Ципфель, П.Ф. (1997) Координация экспрессии и различных характеристик ДНК-связывания четырех белков EGR-цинковых пальцев в Т-лимфоцитах Jurkat. Иммунобиология, 198, 179–191. [PubMed] [Академия Google]
51. Дюран Д.Б., Буш М.Р., Морган Дж.Г., Весс А. и Крэбтри, Г.Р. (1988) Характеристика элементов ответа антигенного рецептора в энхансере интерлейкина-2. Мол. Клетка. биол., 8, 1715–1724. [Бесплатная статья PMC] [PubMed] [Google Scholar]
52. Jain J., McCaffrey, P.G., Valge-Archer, V.E. и Рао, А. (1992) Анализ сайтов AP-1 в промоторе IL-2. Природа, 356, 801–804. [Google Scholar]
53. Цай Э.Ю., Джейн Дж., Песавенто П.А., Рао А. и Голдфельд, А.Е. (1996) Регуляция гена фактора некроза опухоли альфа в активированных Т-клетках включает ATF-2/Jun и NFATp. Мол. Клетка. биол., 16, 459–467. [Бесплатная статья PMC] [PubMed] [Google Scholar]
54. Goldfield AE, McCaffrey, P.G., Stromonger, J.K. и Рао, А. (1993) Идентификация нового чувствительного к циклоспорину элемента в промоторе гена фактора некроза опухоли α человека. Дж. Эксп. Мед., 178, 1365–1379. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Дж. Эксп. Мед., 178, 1365–1379. [Бесплатная статья PMC] [PubMed] [Google Scholar]
55. Russo M.W., Sevetson, B.R. и Милбрандт, Дж. (1995) Идентификация NAB1, репрессора транскрипции, опосредованной NGFI-A и Krox20. проц. Натл акад. науч. США, 92, 6873–6877. [Бесплатная статья PMC] [PubMed] [Google Scholar]
56. Сварен Дж., Севетсон Б.Р., Апель Э.Д., Зимонич Д.Б., Попеску Н.К. и Милбрандт, Дж. (1996) Nab2, корепрессор NGFI-A (Egr-1) и Krox20, индуцируется пролиферативными и дифференцированными стимулами. Мол. Клетка. биол., 16, 3545–3553. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Основы EGR — что они делают, как они работают, как устранять неполадки Клапан рециркуляции, или сокращенно клапан рециркуляции отработавших газов, рециркулирует точно дозированные количества выхлопных газов во впускную систему двигателя для повышения эффективности двигателя, снижения расхода топлива и снижения выбросов NOx.
В связи с растущим давлением на снижение выбросов клапан рециркуляции отработавших газов будет играть все более важную роль в будущем.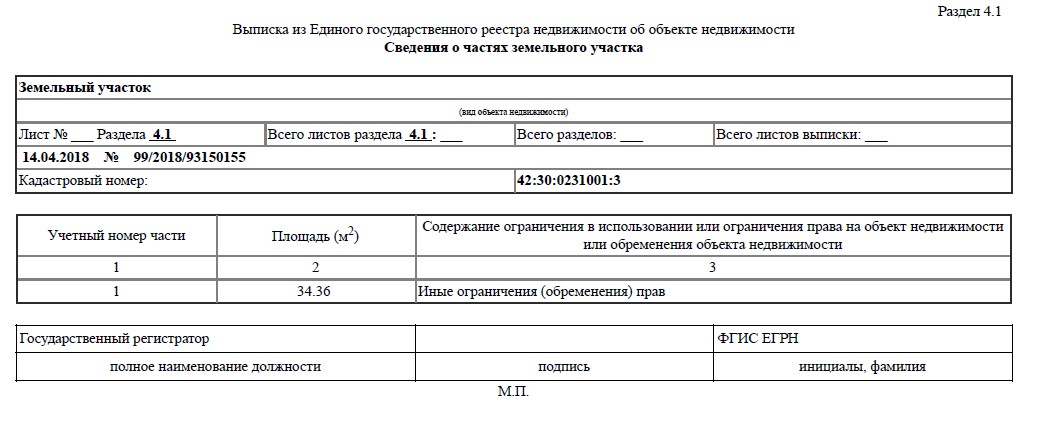 Важно знать, что он делает, почему он выходит из строя и как заменить его, когда он выходит из строя.
Важно знать, что он делает, почему он выходит из строя и как заменить его, когда он выходит из строя.
Как работает клапан EGR?
Примеры агрессивных сред для клапанов EGR.
Почти 80 процентов воздуха, которым мы дышим, состоит из азота. Однако, когда он подвергается воздействию чрезвычайно высоких температур в камере сгорания, плюс 1370°C, обычно инертный газ становится реактивным, создавая вредные оксиды азота или NOx, которые затем выбрасываются через выхлопную систему в атмосферу.
Чтобы свести это к минимуму, клапан рециркуляции отработавших газов позволяет определенному количеству выхлопных газов повторно поступать во впускную систему, эффективно изменяя химический состав воздуха, поступающего в двигатель. С меньшим количеством кислорода теперь разбавленная смесь сгорает медленнее, снижая температуру в камере сгорания почти на 150 ° C и уменьшая образование NOx для более чистого и эффективного выхлопа.
Клапан рециркуляции отработавших газов имеет два основных положения: открытое и закрытое, хотя положение может варьироваться в любом промежуточном положении. Клапан EGR закрыт, когда двигатель запускается. На холостом ходу и на малых оборотах требуется лишь небольшое количество мощности, а следовательно, лишь небольшое количество кислорода, поэтому клапан открывается постепенно – может быть до 90% открыт на холостом ходу. Однако, когда требуется больший крутящий момент и мощность, например, при полном ускорении, клапан рециркуляции отработавших газов закрывается, чтобы обеспечить поступление в цилиндр как можно большего количества кислорода.
Клапан EGR закрыт, когда двигатель запускается. На холостом ходу и на малых оборотах требуется лишь небольшое количество мощности, а следовательно, лишь небольшое количество кислорода, поэтому клапан открывается постепенно – может быть до 90% открыт на холостом ходу. Однако, когда требуется больший крутящий момент и мощность, например, при полном ускорении, клапан рециркуляции отработавших газов закрывается, чтобы обеспечить поступление в цилиндр как можно большего количества кислорода.
Клапаны рециркуляции отработавших газов не только снижают выбросы NOx, но и могут использоваться в двигателях GDi уменьшенных размеров для снижения насосных потерь и повышения эффективности сгорания и устойчивости к детонации. В дизеле это также может помочь уменьшить детонацию дизельного двигателя на холостом ходу.
Типы клапана EGR
Хотя существует несколько типов клапана EGR — в более ранних системах используется клапан с вакуумным приводом, а в новых автомобилях используется электронное управление — основные типы можно в общих чертах свести к следующим:
Дизельные клапаны системы рециркуляции отработавших газов высокого давления отводят поток выхлопных газов с высоким содержанием сажи до того, как они попадут в сажевый фильтр — сажа может смешиваться с парами масла, образуя шлам. Затем газ возвращается во впускной коллектор либо через трубу, либо через внутренние отверстия в головке блока цилиндров. Вторичный клапан также используется для создания вакуума во впускном коллекторе, поскольку он отсутствует в дизельных двигателях.
Дизель Клапаны EGR низкого давления отводят выхлопные газы после их прохождения через сажевый фильтр — этот газ имеет меньший поток, но почти полностью очищен от сажи. Затем газ возвращается во впускной коллектор по трубе.
Бензиновые клапаны рециркуляции отработавших газов отводят отработавшие газы так же, как дизельный аналог высокого давления. Вакуум, создаваемый разрежением цилиндра, втягивает выхлопные газы, а поток регулируется открытием и закрытием самого клапана рециркуляции отработавших газов.
Вакуумные клапаны EGR используют вакуумный соленоид для изменения разрежения на диафрагме и, в свою очередь, открытия и закрытия EGR. Некоторые клапаны также оснащены датчиком обратной связи для информирования ЭБУ о положении клапанов.
Цифровые клапаны EGR оснащены электромагнитным или шаговым двигателем и, в большинстве случаев, датчиком обратной связи. Эти клапаны получают сигнал с широтно-импульсной модуляцией от ЭБУ для регулирования потока выхлопных газов.
Почему выходят из строя клапаны EGR?
Клапаны EGR работают в агрессивной среде, поэтому со временем изнашиваются. Тем не менее, самой большой причиной отказа является накопление частиц углерода из выхлопных газов вдоль каналов EGR и системы впуска. Со временем это засорит трубки, каналы для выхлопных газов и, в конечном итоге, плунжерный механизм клапана, в результате чего он либо заклинит в открытом, либо в закрытом состоянии. Неисправности также могут быть вызваны разрывом или протечкой мембраны клапана.
На что обратить внимание при неисправности клапана рециркуляции отработавших газов?
Симптомы, связанные с отказом клапана EGR, аналогичны симптомам многих других компонентов системы управления двигателем, и из-за этого неисправности EGR продолжают вызывать головную боль у многих техников. Однако есть несколько признаков, на которые следует обратить внимание:
Однако есть несколько признаков, на которые следует обратить внимание:
- Индикатор проверки двигателя: , как и в случае с большинством компонентов управления двигателем, проблема с клапаном рециркуляции отработавших газов может вызвать появление индикатора проверки двигателя.
- Проблемы с работой двигателя: , если клапан застрял в открытом положении, соотношение воздух-топливо автомобиля будет нарушено, что приведет к проблемам с работой двигателя, таким как снижение мощности, плохое ускорение и неровный холостой ход. Это также может привести к утечке давления турбонаддува, из-за чего турбонаддув будет работать тяжелее.
- Повышенный выброс NOx: , когда клапан рециркуляции отработавших газов остается закрытым, возникающие в результате высокие температуры в камере сгорания оставляют много несгоревшего топлива в выхлопных газах, что приводит к увеличению выбросов NOx и снижению эффективности использования топлива.
- Детонация в двигателе: более высокие температуры и NOx также могут привести к усилению детонации или детонации, которые слышны как стук в двигателе.
Поиск и устранение неисправностей клапана рециркуляции отработавших газов
Учитывая различные типы клапанов рециркуляции отработавших газов, всегда лучше следовать процедурам устранения неполадок, подробно описанным в руководстве по обслуживанию, однако есть несколько общих шагов, которые могут помочь в точной диагностике:
- Читать любые коды неисправностей на клапанах EGR с электронным управлением с помощью диагностического прибора.
- Убедитесь, что все вакуумные линии и электрические соединения подсоединены и расположены правильно.
- С помощью вакуумметра проверьте шланг подачи вакуума на наличие вакуума при частоте вращения от 2000 до 2500 об/мин. Отсутствие вакуума при нормальных рабочих температурах указывает на ослабленный шланг, заблокированный или неисправный вакуумный переключатель или соленоид или неисправный вакуумный усилитель/насос.
- Проверьте вакуумный соленоид при работающем двигателе. На клапанах EGR с электронным управлением активируйте соленоид с помощью диагностического прибора и проверьте вакуум на конце трубы. Если соленоид не открывается при подаче питания, застрял в открытом или закрытом положении или имеет корродированное электрическое соединение, ослабленный провод или плохое заземление, это повлияет на работу EGR. Перед заменой определите основную причину.
- Если возможно, проверьте движение штока клапана в диапазоне от 1500 до 2000 об/мин. Шток клапана должен двигаться, если клапан работает правильно, если нет и есть вакуум, значит, есть неисправность.
- Подайте вакуум непосредственно к клапану EGR с помощью ручного вакуумного насоса или диагностического прибора в зависимости от типа клапана EGR. Если нет изменения качества холостого хода, то либо неисправен клапан EGR, либо проходы полностью забиты. Если двигатель работает с перебоями на холостом ходу или глохнет, проблема вызвана неисправностью системы управления.
- Снимите клапан рециркуляции отработавших газов и проверьте его на наличие нагара. По возможности удалите нагар, стараясь не загрязнить диафрагму.
- Осмотрите канал рециркуляции отработавших газов в коллекторе на предмет засорения и при необходимости очистите.


Общие коды неисправностей EGR
На клапанах EGR последних моделей распространены следующие коды неисправностей:
- P0400: Неисправность потока EGR
- P0401: Обнаружен недостаточный поток EGR
- P0402: Обнаружен избыточный поток EGR
- P0403: Неисправность цепи EGR
- P0404: Диапазон/функционирование цепи EGR
- P0405: Низкий уровень сигнала в цепи датчика А системы рециркуляции отработавших газов
- P0406: Высокий уровень сигнала цепи датчика А системы рециркуляции отработавших газов
- P0407: Низкий уровень сигнала в цепи датчика В системы рециркуляции отработавших газов
- P0408: Высокий уровень сигнала в цепи датчика В системы рециркуляции отработавших газов
- P1403: Соленоид EGR низкий уровень
- P1404: Система рециркуляции отработавших газов — ошибка иглы закрытого клапана
- P1405: Высокий уровень соленоида EGR
- P1406: Ошибка положения штифта системы EGR
Как заменить неисправный клапан EGR?
- Сначала снимите кожух двигателя.
- Затем ослабьте электрический кабель на клапане и отсоедините электрические соединения и/или вакуумные линии, проверив наличие признаков повреждения.
- Снимите крепежные винты и проверьте клапан на наличие повреждений, коррозии или нагара.
- Тщательно очистите монтажную поверхность клапана EGR и установите новый клапан и прокладку. Удалите также свободный углерод из порта подачи EGR.
- Совместите клапан рециркуляции отработавших газов с отверстиями для болтов и прокладкой и снова прикрепите к корпусу.
- Затяните все крепежные детали с рекомендованным крутящим моментом.
- После надежной установки снова подсоедините вакуумные линии и/или электрические соединения.
- Наконец, используйте диагностический сканер, чтобы сбросить индикатор управления двигателем и проверить наличие других ошибок. Убедитесь, что контрольная лампа неисправности погасла, затем проведите дорожное испытание. Многие автомобили теперь требуют сброса клапана рециркуляции отработавших газов в соответствии с адаптацией. Это просто позволяет ECU узнать положение остановки в точках открытия и закрытия. Если этого не сделать, клапан может сломаться и упасть в коллектор.

 Это просто позволяет ECU узнать положение остановки в точках открытия и закрытия. Если этого не сделать, клапан может сломаться и упасть в коллектор.
Это просто позволяет ECU узнать положение остановки в точках открытия и закрытия. Если этого не сделать, клапан может сломаться и упасть в коллектор.Огромная дополнительная добыча ископаемого топлива и рост выбросов, связанных с климатом, в результате геоинженерных проектов в Азии — Geoengineering Monitor
См. в формате PDF
Автор: Аня Чалмин
Введение
В Азии количество новых и планируемых улавливания и хранения углерода ( CCS) проектов быстро увеличилось в последние годы. Захваченный CO 2 в основном используется для увеличения добычи нефти (EOR) или увеличения добычи газа (EGR). EOR и EGR включают перекачку CO 9 под давлением0265 2 в нефтяные или газовые резервуары для извлечения оставшихся запасов из стареющих нефтяных и газовых месторождений, а также для добычи ископаемых видов топлива, недоступных другим способом, что значительно увеличивает добычу. Эта технология была разработана нефтяной промышленностью полвека назад для освоения труднодоступных глубинных запасов нефти, и теперь она продается под новым названием и с оттенком зелени.
Взглянув на один пример, балансы выбросов CO 2 китайских проектов Sinopec CCS, становится ясно, что CCS не направлен на сокращение выбросов CO 2 выбросов, как нам и полагается считать, а только об увеличении объемов добычи ископаемых. Проекты утверждают, что хранят CO 2 , но на самом деле приводят к множеству источников новых выбросов, например, выбросы CO 2 на электростанции Shengli увеличились более чем в пять раз из-за проекта CCS. Поскольку произведенные выбросы CO 2 могут быть рассчитаны лишь частично, в действительности результат, вероятно, будет еще более разрушительным.
Выбросы от добычи ископаемого топлива, вероятно, продолжат увеличиваться, так как нефтяные и газовые месторождения становятся все более труднодоступными и расположены на все большей глубине – это означает, например, более длинные расстояния перекачки и еще большие объемы воды для выталкивания сырья на поверхность с глубины в несколько километров.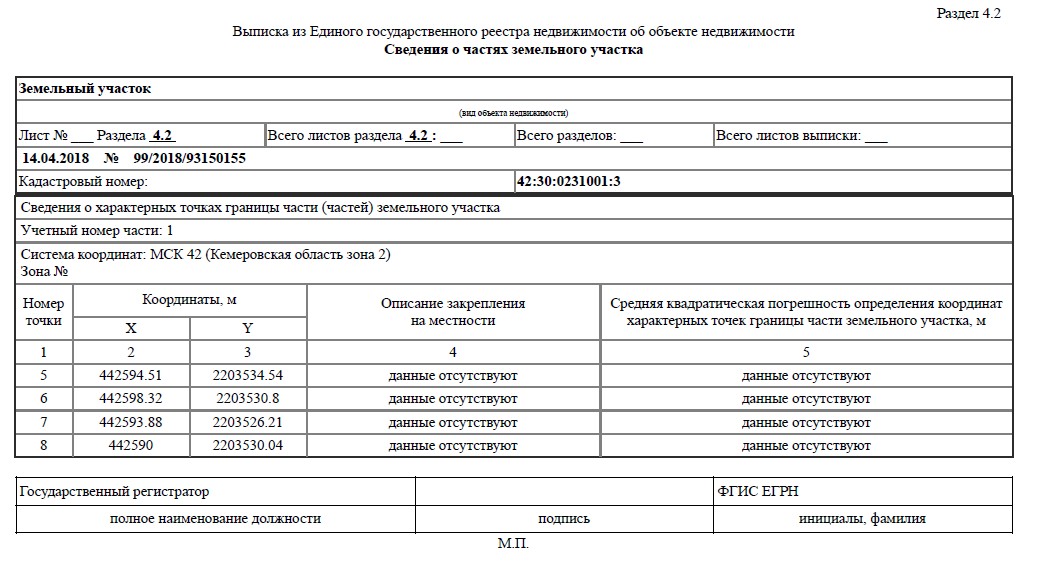
В некоторых регионах Азии промышленность развивается вокруг использования уловленного CO 2 , например, для производства химикатов или газированных напитков. Однако должно быть ясно, с одной стороны, что это энергоемкие этапы переработки, которые генерируют выбросы, а с другой стороны, что это не долговременное хранение СО 2 , так как переработанный СО 2 высвобождается снова после того, как продукты потребляются.
Повышение нефтеотдачи не только увеличивает выбросы CO
2 , но и увеличивает их
В Азии улавливаемый CO 2 в основном используется для EOR и EGR. В этом разделе объясняются катастрофические климатические последствия этой практики с помощью некоторых конкретных количественных примеров. Цифры показывают: EOR и EGR увеличивают выбросы CO 2 , даже по сравнению с количеством захваченного CO 2 . Другие выбросы и затраты, такие как энергия, необходимая для улавливания CO 2 , выбросы метана на объектах добычи, затраты на здоровье и окружающую среду, связанные с добычей и загрязняющими веществами, не могли быть приняты во внимание из-за отсутствия данных.
В своем годовом отчете за 2021 год Sinopec, вторая по величине энергетическая компания Китая, сообщает об использовании 0,31 миллиона тонн уловленного CO 2 для увеличения нефтеотдачи в 2021 году и отмечает в дополнительном выпуске, что эти действия по увеличению нефтеотдачи увеличили добычу нефти на 0,089 миллиона тонн. Агентство по охране окружающей среды США (EPA) дает в среднем 0,43 тонны выбросов CO 2 на баррель сырой нефти, а Норвежское нефтяное управление (NPD) заявляет, что одна тонна сырой нефти соответствует 7,49бочки. Таким образом, одна тонна сырой нефти эквивалентна примерно 3,22 тоннам выбросов CO 2 . При выбросах 0,43 тонны CO 2 на баррель сырой нефти увеличение добычи нефти на 0,089 миллиона тонн сырой нефти приводит к дополнительным выбросам 0,29 миллиона тонн CO 2 . Эта сумма не включает следующие дополнительные выбросы и затраты CO 2 :
- Процесс улавливания CO 2 на электростанции требует больших затрат энергии, что увеличивает потребление ископаемого топлива до 30 %, т. е. , необходимо добывать и сжигать еще больше ископаемых энергетических ресурсов, чтобы производить такое же количество энергии с помощью CCS.
- Около 30 % уловленного CO 2 , используемого для МУН, немедленно повторно попадает в атмосферу. Это соответствует 0,093 миллиона тонн выбросов CO 2 в приведенном выше примере.
- При добыче и транспортировке сырой нефти образуются выбросы, в среднем 95 кг эквивалента CO 2 на баррель сырой нефти, что соответствует 0,7 тонны эквивалента CO 2 на тонну сырой нефти. Это означает дополнительно 0,062 миллиона тонн CO 2 эквивалентные выбросы для примера, рассчитанного выше.
- На добывающих предприятиях происходят климатически значимые выбросы метана.
- Сжигание ископаемого сырья приводит к высоким затратам на окружающую среду и здоровье людей из-за содержащихся в нем загрязняющих веществ.
 е. , необходимо добывать и сжигать еще больше ископаемых энергетических ресурсов, чтобы производить такое же количество энергии с помощью CCS.
е. , необходимо добывать и сжигать еще больше ископаемых энергетических ресурсов, чтобы производить такое же количество энергии с помощью CCS. Даже если не учитывать значительные дополнительные выбросы в результате энергоемкого процесса улавливания CO 2 и вызванные выбросы метана, описанный выше пример CCS-EOR вызывает больше выбросов CO 2 (0,445 миллиона тонн), чем CO 2 улавливается (0,31 млн тонн). Кроме того, нет никакой гарантии, что CO 2 , сжатый под землей, не попадет обратно в атмосферу через одну из многочисленных скважин в подземных газовых месторождениях, в которые закачивается CO 2 , или через тектонические движения.
Кроме того, нет никакой гарантии, что CO 2 , сжатый под землей, не попадет обратно в атмосферу через одну из многочисленных скважин в подземных газовых месторождениях, в которые закачивается CO 2 , или через тектонические движения.
В январе 2022 года Sinopec завершила модернизацию своего завода по производству удобрений Qilu Petrochemical в городе Цзыбо в провинции Шаньдун с помощью системы улавливания CO 2 и объявила, что улавливаемый CO 2 будет транспортироваться по трубопроводу на нефтяное месторождение Шэнли для повышения нефтеотдачи. Ожидается, что установка улавливания CO 2 на этом участке будет введена в эксплуатацию в 2022 году. На своем веб-сайте Sinopec заявила: « Ожидается, что проект также увеличит добычу сырой нефти почти на 3 миллиона тонн в следующие 15 лет». Это подтверждается сообщениями прессы: « Sinopec рассчитывает повторно закачать 10,68 млн тонн CO 2 в скважины в течение следующих 15 лет и иметь возможность увеличить добычу нефти в общей сложности на 22 миллиона баррелей ». Этот дополнительный объем производства сырой нефти в течение следующих 15 лет соответствует 9,6 миллионам тонн дополнительных выбросов CO 2 . С учетом выбросов, образующихся при добыче и транспортировке (в среднем 95 кг CO 2 эквивалента на баррель сырой нефти или 0,7 тонны CO 2 эквивалента на тонну сырой нефти), это соответствует еще 2,1 млн. тонн CO 2 эквивалента – всего 11,7 млн тонн дополнительного CO 2 выбросы. Опять же, это не принимает во внимание энергоемкую технологию улавливания CO 2 , вызванные выбросы метана, затраты на окружающую среду и здоровье, а также CO 2 , возвращающийся в атмосферу.
Этот дополнительный объем производства сырой нефти в течение следующих 15 лет соответствует 9,6 миллионам тонн дополнительных выбросов CO 2 . С учетом выбросов, образующихся при добыче и транспортировке (в среднем 95 кг CO 2 эквивалента на баррель сырой нефти или 0,7 тонны CO 2 эквивалента на тонну сырой нефти), это соответствует еще 2,1 млн. тонн CO 2 эквивалента – всего 11,7 млн тонн дополнительного CO 2 выбросы. Опять же, это не принимает во внимание энергоемкую технологию улавливания CO 2 , вызванные выбросы метана, затраты на окружающую среду и здоровье, а также CO 2 , возвращающийся в атмосферу.
Sinopec также является владельцем и оператором электростанции Shengli в городе Дунъин провинции Шаньдун. CO 2 с 2010 года улавливается старым блоком электростанции, работающим на угле. Количество составляет до 36 500 тонн CO 9.0265 2 в год. Захваченный CO 2 транспортируется автоцистерной примерно на 80 километров на нефтяное месторождение Шэнли для повышения нефтеотдачи. Сколько дополнительной сырой нефти было добыто с этим количеством CO 2 , не раскрывается, но если применить среднее значение деятельности Sinopec по увеличению нефтеотдачи в 2021 году, оно составит 10 489 тонн сырой нефти с выбросом 33 773 тонн CO 2 . Это количество CO 2 еще не включает следующие дополнительные выбросы CO 2 :
Сколько дополнительной сырой нефти было добыто с этим количеством CO 2 , не раскрывается, но если применить среднее значение деятельности Sinopec по увеличению нефтеотдачи в 2021 году, оно составит 10 489 тонн сырой нефти с выбросом 33 773 тонн CO 2 . Это количество CO 2 еще не включает следующие дополнительные выбросы CO 2 :
- Процесс улавливания CO 2 на угольной электростанции требует больших энергозатрат, что увеличивает потребление ископаемого топлива до 30 %, т. е. для получения того же количества необходимо добыть и сжечь еще больше угля. энергии с CCS.
- Перевозка автоцистерной на расстояние около 80 км для МУН: при среднем контрольном выбросе 52,7 граммов CO 2 на тонно-километр перевозка 36 500 тонн СО 2 на расстояние более 80 км для МУН соответствует 4 тоннам 153,88. СО 2 выбросы.
- Около 30 % уловленного CO 2 , используемого для МУН, будет напрямую выбрасываться обратно в атмосферу, что соответствует 10 950 тоннам CO 2 в год.
- Добыча и транспортировка сырой нефти: В среднем 95 кг эквивалента CO 2 на баррель сырой нефти, что соответствует 0,7 тонны эквивалента CO 2 на тонну сырой нефти. Это означает 7 342 тонны дополнительных выбросов CO 2 эквивалента.

Несмотря на значительные дополнительные выбросы в результате энергоемкого процесса улавливания CO 2 на угольной электростанции, проект CCS-EOR в Шэнли по-прежнему производит в пять раз больше выбросов CO 2 (205 949 тонн) чем улавливается CO 2 (36 500 тонн).
В течение этого десятилетия Sinopec планирует увеличить доступную мощность по улавливанию CO 2 на электростанции Shengli до одного миллиона тонн CO 2 в год. Эти планы уже предполагалось реализовать в 2017 году, причина переноса не разглашается.
Согласно базе данных CCS и прессе, мероприятия по увеличению нефтеотдачи увеличили добычу нефти на нефтяном месторождении Шэнли, одном из крупнейших и старейших нефтяных месторождений в Китае, на 10–15 %. Согласно базе данных CCS Массачусетского технологического института, годовая добыча нефтяного месторождения Шэнли составила 27 миллионов тонн нефти в 2012 году, через два года после начала работ по CCS-EOR. Увеличение от 10 % до 15 % с МУН до 27 миллионов тонн соответствует от 2,45 миллиона тонн до 3,5 миллиона тонн сырой нефти. Это увеличение, конечно, не было достигнуто с улавливанием 36 500 тонн уловленного CO 9.0265 2 -EOR, но этот сценарий, тем не менее, следует рассчитать, чтобы проиллюстрировать влияние крупномасштабных мероприятий EOR. Применяя приведенные выше значения EPA и NPD (средние) США, это означает, что ежегодно образуется от 7,89 млн тонн (10%) до 11,27 млн тонн (15%) дополнительных выбросов CO 2 .
Согласно базе данных CCS Массачусетского технологического института, годовая добыча нефтяного месторождения Шэнли составила 27 миллионов тонн нефти в 2012 году, через два года после начала работ по CCS-EOR. Увеличение от 10 % до 15 % с МУН до 27 миллионов тонн соответствует от 2,45 миллиона тонн до 3,5 миллиона тонн сырой нефти. Это увеличение, конечно, не было достигнуто с улавливанием 36 500 тонн уловленного CO 9.0265 2 -EOR, но этот сценарий, тем не менее, следует рассчитать, чтобы проиллюстрировать влияние крупномасштабных мероприятий EOR. Применяя приведенные выше значения EPA и NPD (средние) США, это означает, что ежегодно образуется от 7,89 млн тонн (10%) до 11,27 млн тонн (15%) дополнительных выбросов CO 2 .
Проект Sinopec Eastern China CCS расположен на заводе Sinopec Nanjing Chemical Industries Co. Ltd. в городе Нанкин, провинция Цзянсу. Завод выбрасывает CO 2 в производстве синтетического аммиака и при переработке угля в водород. Демонстрационная установка для улавливания 0,05 млн тонн CO 2 была введена в эксплуатацию на площадке в 2015 году, а уловленный CO 2 используется для увеличения нефтеотдачи на близлежащих нефтяных месторождениях. Уже в 2019 году Sinopec объявила о планах по реализации более крупного проекта CCS на заводе. Завершение проекта первоначально было объявлено на 2021 год, но затем было перенесено на 2025 год. По сообщениям прессы, новая установка улавливания будет рассчитана на улавливание 0,2 миллиона тонн CO 9.0265 2 в год и, как ожидается, « увеличит добычу на месторождении на 1320 баррелей в день ». Это эквивалентно 64 326 тоннам сырой нефти в год и соответствует 207 129 тоннам выбросов CO 2 – без учета дополнительных выбросов от CCS-EOR, упомянутых выше.
Уже в 2019 году Sinopec объявила о планах по реализации более крупного проекта CCS на заводе. Завершение проекта первоначально было объявлено на 2021 год, но затем было перенесено на 2025 год. По сообщениям прессы, новая установка улавливания будет рассчитана на улавливание 0,2 миллиона тонн CO 9.0265 2 в год и, как ожидается, « увеличит добычу на месторождении на 1320 баррелей в день ». Это эквивалентно 64 326 тоннам сырой нефти в год и соответствует 207 129 тоннам выбросов CO 2 – без учета дополнительных выбросов от CCS-EOR, упомянутых выше.
Осенью 2021 года Sinopec объявила об открытии новых запасов нефти на своем месторождении Шэнли, включающих 458 миллионов тонн сланцевого масла и дебит до 0,062 миллиона тонн сланцевого масла в год. В отчете, подготовленном для Немецкой ассоциации возобновляемых источников энергии, сравниваются выбросы CO 2 выбросы различных ископаемых видов топлива и описывает примерно на 40 % более высокие выбросы при использовании сланцевой нефти по сравнению с сырой нефтью (см. рис. 2) из-за состава сланцевой нефти, более длинных расстояний перекачки и повышенного водонефтяного отношения. Это означает, что на тонну сланцевого масла выбрасывается 4,5 тонны CO 2 , что соответствует 2,061 миллиарда тонн CO 2 по отношению к недавно открытому месторождению сланцевой нефти на месторождении Шэнли. Это лишь одно из многих месторождений, а Шэнли — одно из нескольких месторождений нефти и газа, где Sinopec ведет добычу ископаемого топлива. Шкала СО 2 выбросы, связанные с этим единственным объектом, должны показать, насколько неэффективен CCS, даже после десятилетий разработки и обширного финансирования исследований, даже если бы для захваченного CO 2 существовало безопасное хранилище. Общее количество CO 2 , захваченное Sinopec в 2021 году (0,31 миллиона тонн), эквивалентно 0,00015 % содержания CO 2 в этом единственном недавно открытом запасе нефти.
рис. 2) из-за состава сланцевой нефти, более длинных расстояний перекачки и повышенного водонефтяного отношения. Это означает, что на тонну сланцевого масла выбрасывается 4,5 тонны CO 2 , что соответствует 2,061 миллиарда тонн CO 2 по отношению к недавно открытому месторождению сланцевой нефти на месторождении Шэнли. Это лишь одно из многих месторождений, а Шэнли — одно из нескольких месторождений нефти и газа, где Sinopec ведет добычу ископаемого топлива. Шкала СО 2 выбросы, связанные с этим единственным объектом, должны показать, насколько неэффективен CCS, даже после десятилетий разработки и обширного финансирования исследований, даже если бы для захваченного CO 2 существовало безопасное хранилище. Общее количество CO 2 , захваченное Sinopec в 2021 году (0,31 миллиона тонн), эквивалентно 0,00015 % содержания CO 2 в этом единственном недавно открытом запасе нефти.
Большое количество проектов CCS находится в стадии реализации
В этом разделе представлен обзор проектов CCS в Азии и на Ближнем Востоке. Из-за большого количества проектов в разделе основное внимание уделяется новым разработкам, запланированным проектам и крупным текущим проектам. Интерактивная карта мира по геоинженерии, подготовленная ETC Group и Фондом Генриха Бёлля, дает представление о дальнейших проектах. В большинстве новых и текущих проектов планируется развертывание CCS для увеличения добычи нефти (EOR) или увеличения добычи газа (EGR).
Из-за большого количества проектов в разделе основное внимание уделяется новым разработкам, запланированным проектам и крупным текущим проектам. Интерактивная карта мира по геоинженерии, подготовленная ETC Group и Фондом Генриха Бёлля, дает представление о дальнейших проектах. В большинстве новых и текущих проектов планируется развертывание CCS для увеличения добычи нефти (EOR) или увеличения добычи газа (EGR).
Бахрейн
В январе 2022 года Mitsubishi Heavy Industries EMEA, Ltd., разработчик технологии улавливания CO 2 , и Aluminium Bahrain B.S.C. (Alba), крупный производитель алюминия в Бахрейне, подписал меморандум о взаимопонимании для изучения возможности улавливания CO 2 из выхлопных газов алюминиевого завода, которым управляет Alba в Бахрейне. Информация о сроках реализации проекта, планируемом количестве уловленного CO 2 и использовании уловленного CO 2 остаются нераскрытыми.
Китай
Британско-китайский центр CCUS в провинции Гуандун (GDCCUSC) был создан для тестирования и продвижения CCS и CCUS в Китае и финансируется государственными органами Великобритании и Китая. GDCCUSC выбрал угольную электростанцию Haifeng для тестирования пяти различных технологий улавливания CO 2 , включая два метода на основе аминов, два метода на основе мембран и один метод физического поглощения. Крупномасштабный проект CCS в Хайфэне с запланированным ежегодным CO 2 мощность улавливания один миллион тонн, планируется после завершения испытаний технологии улавливания CO 2 . Геологическое морское хранилище CO 2 для проекта Хайфэн исследуется в близлежащем бассейне устья Жемчужной реки. Согласно предварительному анализу, бассейн подходит для МУН, «, но технико-экономическое обоснование требует дополнительной оценки ». Ввод в эксплуатацию более крупного ЦО 2 9Проект улавливания 0266 первоначально был объявлен на 2020-е годы, но в этом году он был перенесен на 2030 год. угольная электростанция в деревне Чжуозитоу, уезд Чжэннин, провинция Ганьсу. Планируется, что проект будет запущен к 2023 году, и уловленный CO 2 будет транспортироваться по трубопроводу для подземного сброса или использования EOR.
GDCCUSC выбрал угольную электростанцию Haifeng для тестирования пяти различных технологий улавливания CO 2 , включая два метода на основе аминов, два метода на основе мембран и один метод физического поглощения. Крупномасштабный проект CCS в Хайфэне с запланированным ежегодным CO 2 мощность улавливания один миллион тонн, планируется после завершения испытаний технологии улавливания CO 2 . Геологическое морское хранилище CO 2 для проекта Хайфэн исследуется в близлежащем бассейне устья Жемчужной реки. Согласно предварительному анализу, бассейн подходит для МУН, «, но технико-экономическое обоснование требует дополнительной оценки ». Ввод в эксплуатацию более крупного ЦО 2 9Проект улавливания 0266 первоначально был объявлен на 2020-е годы, но в этом году он был перенесен на 2030 год. угольная электростанция в деревне Чжуозитоу, уезд Чжэннин, провинция Ганьсу. Планируется, что проект будет запущен к 2023 году, и уловленный CO 2 будет транспортироваться по трубопроводу для подземного сброса или использования EOR. Тяньцзинь, недалеко от Пекина, является домом для демонстрационного проекта GreenGen IGCC группы Huaneng, целью которого является разработка комбинированного цикла интегрированной газификации (IGCC) с CCS. Строительство первой демонстрационной очереди завершено в 2012 г. С 2016 г. дополнительно СО 2 мощности улавливания добавлены, ввод в эксплуатацию запланирован на 2020-е годы. Ожидается, что после завершения проект будет улавливать до 0,1 миллиона тонн CO 2 в год. В настоящее время рассматривается третий этап, который будет включать строительство и эксплуатацию электростанции IGCC мощностью 400 мегаватт с соответствующими установками по улавливанию CO 2 и МУН.
Тяньцзинь, недалеко от Пекина, является домом для демонстрационного проекта GreenGen IGCC группы Huaneng, целью которого является разработка комбинированного цикла интегрированной газификации (IGCC) с CCS. Строительство первой демонстрационной очереди завершено в 2012 г. С 2016 г. дополнительно СО 2 мощности улавливания добавлены, ввод в эксплуатацию запланирован на 2020-е годы. Ожидается, что после завершения проект будет улавливать до 0,1 миллиона тонн CO 2 в год. В настоящее время рассматривается третий этап, который будет включать строительство и эксплуатацию электростанции IGCC мощностью 400 мегаватт с соответствующими установками по улавливанию CO 2 и МУН.
С 2006 года компания Alstom реализует демонстрационный проект CCS на своей угольной электростанции Харбин в провинции Хэйлунцзян. Проект улавливает до 0,2 млн тонн CO 2 в год, а захваченный CO 2 используется для повышения нефтеотдачи на близлежащих нефтяных месторождениях, таких как нефтяное месторождение Дацин. Alstom планировала дополнительный проект CCS в провинции Хэйлунцзян, где 700-мегаваттная угольная электростанция должна была быть введена в эксплуатацию китайской корпорацией Tatang для работ по повышению нефтеотдачи, но проект был отменен.
Alstom планировала дополнительный проект CCS в провинции Хэйлунцзян, где 700-мегаваттная угольная электростанция должна была быть введена в эксплуатацию китайской корпорацией Tatang для работ по повышению нефтеотдачи, но проект был отменен.
С 2017 года компания Ningxia Coal Industry Group Co. Ltd. проводит оценку проекта CCS на своем проекте Shenhua Ningxia Coal-to-Liquid (CTL) в провинции Нинся. Разработка концепции CCS все еще находится на начальной стадии. Использование CO 2 для EOR рассматривается.
Sinopec управляет пилотным и демонстрационным заводом по улавливанию CO 2 на Zhongyuan Petrochemical Co. Ltd., крупном китайском нефтехимическом предприятии, расположенном в Пуяне, провинция Хэнань. Компания является совместным предприятием Sinopec и провинции Хэнань и управляет нефтехимической производственной линией на основе угля, например, для производства аммиака. Захваченный CO 2 объемом до 0,12 млн тонн в год транспортируется на нефтяные месторождения Чжунъюань для повышения нефтеотдачи.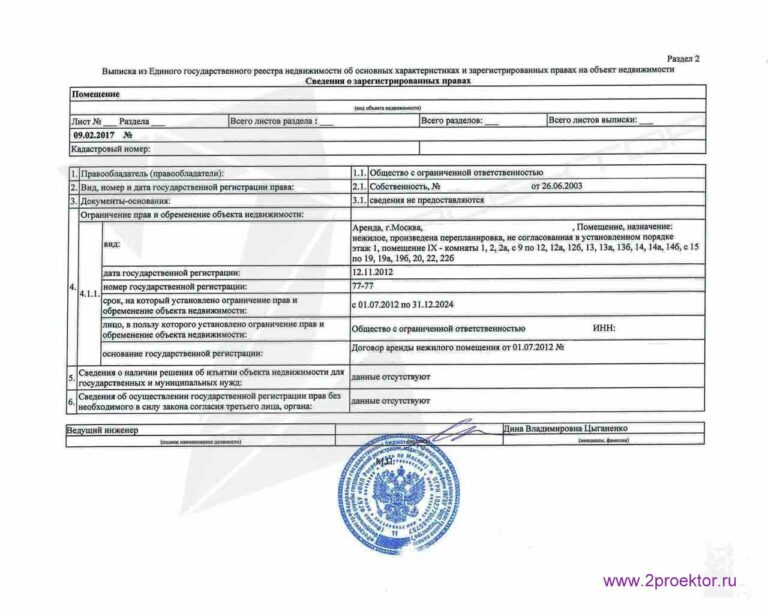 Добыча нефти на месторождениях снижается, и МУН используется для компенсации падения добычи.
Добыча нефти на месторождениях снижается, и МУН используется для компенсации падения добычи.
В 2014 году Shanxi Coal International Energy Group предложила построить угольную электростанцию мощностью 700 мегаватт с CCS в городе Синьчжоу округа Хэцю провинции Шаньси. Первоначально проект планировалось запустить в 2017 году, но он все еще находится на ранней стадии разработки, например, инициаторы изучают возможные маршруты транспортировки CO 2 и места захоронения.
Университет Хуажонг в городе Инчэн в сотрудничестве с отраслевыми партнерами планировал построить демонстрационный завод CCS в провинции Хубэй. СО 2 должен был быть захвачен в 35-мегаваттном кислородно-топливном котле на электростанции Jiuda Salt в Инчэне и использован для увеличения нефтеотдачи. Электростанция была введена в эксплуатацию в 2015 году. По данным Global CCS Institute, ввод в эксплуатацию компонента CCS планировался на 2020-е годы. В марте 2022 года сообщалось, что работа над проектом CCS остановлена. Причина такого решения пока не обнародована.
Причина такого решения пока не обнародована.
Угольная электростанция Guodian Taizhou принадлежит и управляется компанией Guodian Taizhou Power Generation и расположена в уезде Гаоган, префектура Тайчжоу, провинция Цзянсу. Оператор планирует CO 2 проект по улавливанию, запуск которого ожидается в 2023 году. Планируемая мощность по улавливанию составляет 0,5 миллиона тонн CO 2 , из которых 0,3 миллиона тонн в год будет транспортироваться в Sinopec East China Petroleum для увеличения нефтеотдачи.
В 2021 году Китайская национальная морская нефтяная корпорация (CNOOC) запустила морской проект CCS в Южно-Китайском море, закачивая 0,3 миллиона тонн CO 2 в год в морские резервуары на глубине 80 метров. Проект расположен на нефтяном месторождении Энпин 15-1 в устье Жемчужной реки, около 190 км к юго-востоку от Гонконга.
Индия
С 2019 года Индийская нефтегазовая корпорация (ONGC) и Индийская нефтяная корпорация (IOCL) изучают проект по улавливанию CO 2 на нефтеперерабатывающем заводе IOCL в Кояли в индийском штате Гуджарат. Партнеры по проекту намерены использовать захваченный CO 2 для повышения нефтеотдачи на соседнем нефтяном месторождении Гандхар. В 2021 году компания Dastur International из США получила контракт на проектирование и технико-экономическое обоснование проекта. В апреле 2022 года ONGC подписала меморандум о взаимопонимании с норвежской компанией Equinor ASA о сотрудничестве в реализации проекта.
Партнеры по проекту намерены использовать захваченный CO 2 для повышения нефтеотдачи на соседнем нефтяном месторождении Гандхар. В 2021 году компания Dastur International из США получила контракт на проектирование и технико-экономическое обоснование проекта. В апреле 2022 года ONGC подписала меморандум о взаимопонимании с норвежской компанией Equinor ASA о сотрудничестве в реализации проекта.
Малайзия
Малазийская государственная компания Petronas намеревается разработать проект CCS на газовом месторождении Касавари у побережья Саравака, Малайзия. Это будет включать переработку газа с газового месторождения Касавари на комплексе Petronas LNG в Бинтулу, Малайзия. Завод по переработке СПГ будет улавливать CO 2 и закачивать его в истощенное газовое месторождение. Это кислый газ, ископаемый газ со значительным содержанием сероводорода и большим количеством CO 2 . Petronas планирует начать добычу газа в 2023 году, а улавливание и закачку CO 2 — в 2025 году.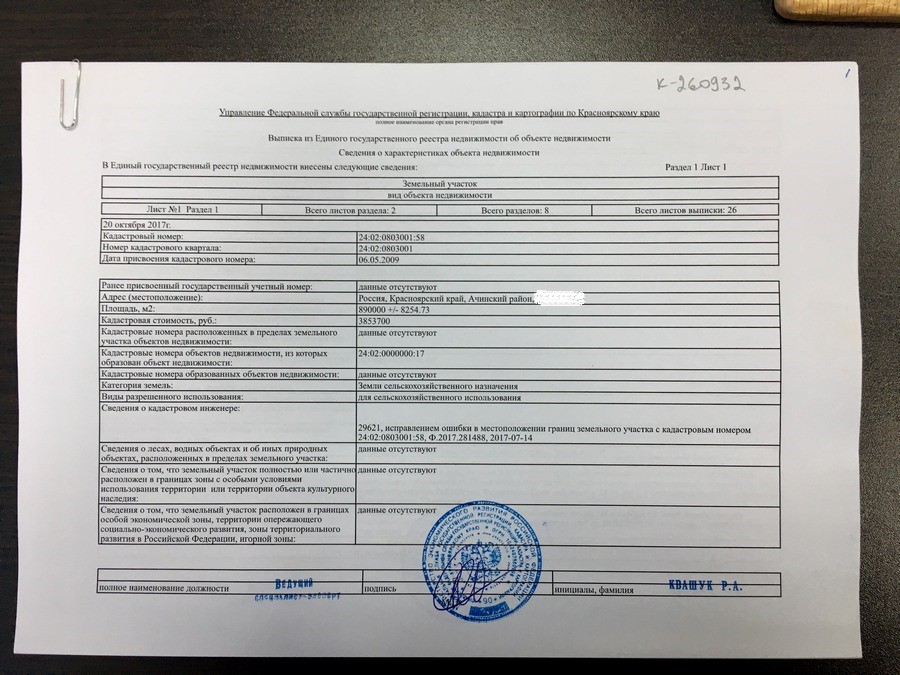 Для финансирования проекта Petronas намерена предложить CCS другим отраслям, таким как энергетика, цемент, сталь и химическая промышленность. В конце 2021 года Petronas подписала меморандумы о взаимопонимании с нефтяной компанией ExxonMobil и производителем стали POSCO.
Для финансирования проекта Petronas намерена предложить CCS другим отраслям, таким как энергетика, цемент, сталь и химическая промышленность. В конце 2021 года Petronas подписала меморандумы о взаимопонимании с нефтяной компанией ExxonMobil и производителем стали POSCO.
Малайзийская нефтегазовая компания Petronas и японская судоходная компания Mitsui OSK. Lines подписали соглашение о совместном изучении возможности транспортировки сжиженного CO 2 в регионах Азиатско-Тихоокеанского региона и Океании.
Qatar
Qatar Energy намеревается построить завод по производству сжиженного природного газа (СПГ) с улавливанием CO 2 в Рас-Лаффане и планирует улавливать до 30% выбросов CO 2 проекта СПГ и выбрасывать их в солевой водоносный горизонт. Total Energies владеет 25% долей в проекте с июня 2022 года.
Россия
Российский производитель газа «Новатэк» намерен разработать проект CCS на заводе по производству сжиженного природного газа (СПГ) «Ямал» в Сабетте на северо-востоке полуострова Ямал. Помимо завода по производству СПГ, проект также включает производство на Южно-Тамбейском газовом месторождении. Новатэк уже выделил две площадки для закачки уловленного CO 2 , Обский на полуострове Ямал и Тадебяяхский на Гыданском полуострове, в каждый из которых, по заявлению компании, будет закачано не менее 600 миллионов тонн СО 2 . В настоящее время рассматривается третье место захоронения. В феврале 2022 года было завершено предварительное инженерно-конструкторское исследование (FEED). «Новатэк» также заявил о планах по поставке голубого водорода, соответствующего европейским стандартам по выбросам CO 2 . «Новатэк» работает над несколькими проектами СПГ в Западной Сибири и стремится увеличить ежегодный экспорт СПГ до 70 миллионов тонн к 2030 году и, в конечном итоге, до более чем 120 миллионов тонн. Ямал СПГ в настоящее время производит 2,6 тонны CO 2 за тонну СПГ.
Помимо завода по производству СПГ, проект также включает производство на Южно-Тамбейском газовом месторождении. Новатэк уже выделил две площадки для закачки уловленного CO 2 , Обский на полуострове Ямал и Тадебяяхский на Гыданском полуострове, в каждый из которых, по заявлению компании, будет закачано не менее 600 миллионов тонн СО 2 . В настоящее время рассматривается третье место захоронения. В феврале 2022 года было завершено предварительное инженерно-конструкторское исследование (FEED). «Новатэк» также заявил о планах по поставке голубого водорода, соответствующего европейским стандартам по выбросам CO 2 . «Новатэк» работает над несколькими проектами СПГ в Западной Сибири и стремится увеличить ежегодный экспорт СПГ до 70 миллионов тонн к 2030 году и, в конечном итоге, до более чем 120 миллионов тонн. Ямал СПГ в настоящее время производит 2,6 тонны CO 2 за тонну СПГ.
Саудовская Аравия
Демонстрационный проект Uthmaniyah CO 2 -EOR управляется государственной нефтяной компанией Saudi Aramco в Королевстве Саудовская Аравия. Проект улавливает 0,8 миллиона тонн CO 2 ежегодно с 2015 года на электростанции Хавия, предприятии по переработке природного газа в Восточной провинции Саудовской Аравии. Захваченный CO 2 транспортируется по трубопроводу длиной 85 километров к береговой точке закачки на производственной установке Утмания на нефтяном месторождении Гавар для повышения нефтеотдачи или используется для производства химикатов.
Проект улавливает 0,8 миллиона тонн CO 2 ежегодно с 2015 года на электростанции Хавия, предприятии по переработке природного газа в Восточной провинции Саудовской Аравии. Захваченный CO 2 транспортируется по трубопроводу длиной 85 километров к береговой точке закачки на производственной установке Утмания на нефтяном месторождении Гавар для повышения нефтеотдачи или используется для производства химикатов.
Южная Корея. улавливается на промышленных предприятиях в юго-восточном городе Ульсан и с 2025 года закачивается в газовый резервуар Тонхэ-1. Резервуар расположен в Японском море, у побережья Южной Кореи. В проекте уже участвуют восемь корейских компаний, в том числе Корейская национальная нефтяная компания в качестве руководителя проекта, SK Energy для улавливания CO 9.0265 2 от промышленных предприятий, SK для переработки CO 2 , Korea Shipbuilding для транспортировки и хранения и Korea Petroleum Corporation, которая эксплуатирует газовое месторождение Тонхэ в течение 20 лет, для закачки CO 2 .
Тимор-Лешти
В июне 2021 года Santos и Eni объявили о совместных планах по созданию хранилища CCS на месторождении Баю-Ундан в Тиморском море, у побережья Тимора-Лешти. Месторождение Баю-Ундан поставляет газ на Дарвинскую электростанцию и расположено примерно в 500 км к северо-западу от Дарвина. Компании, работающие на ископаемом топливе, намерены транспортировать уловленный CO 2 через свой СПГ-терминал в Дарвине, Уикхем-Пойнт, Северная территория, Австралия. CO 2 от их собственной деятельности и, возможно, от других источников выбросов должен быть уловлен, сжат, транспортирован и сброшен на месторождение Баю-Ундан. В марте 2022 года проект вступил в этап начального проектирования и проектирования – этап между технико-экономическими обоснованиями и этапом проектирования, закупок и строительства.
Объединенные Арабские Эмираты
Проект CCS в Абу-Даби — это совместное предприятие компании Abu Dhabi Future Energy Company (Masdar), Emirates Steel Industries (ESI) и Национальной нефтяной компании Абу-Даби (ADNOC). С 2016 года металлургический завод ESI в Муссафе уловил до 0,8 миллиона тонн CO 9.0265 2 как побочный продукт производства чугуна. Захваченный CO 2 затем транспортируется по 50-километровому трубопроводу на нефтяное месторождение Румайта ADNOC, где он используется для увеличения нефтеотдачи. В 2020 году ADNOC объявила о планах увеличить свои текущие мощности по улавливанию CO 2 на 500 %, чтобы к 2030 году достичь пяти миллионов тонн CO 2 в год. местонахождение которого пока не сообщается. Однако в декабре 2021 года ADNOC заключила контракт с инженерно-технологической компанией Technip Energies на разработку неиспользованных запасов нефти и газа на концессионных месторождениях Гаша, включая CCS в разработку.
С 2016 года металлургический завод ESI в Муссафе уловил до 0,8 миллиона тонн CO 9.0265 2 как побочный продукт производства чугуна. Захваченный CO 2 затем транспортируется по 50-километровому трубопроводу на нефтяное месторождение Румайта ADNOC, где он используется для увеличения нефтеотдачи. В 2020 году ADNOC объявила о планах увеличить свои текущие мощности по улавливанию CO 2 на 500 %, чтобы к 2030 году достичь пяти миллионов тонн CO 2 в год. местонахождение которого пока не сообщается. Однако в декабре 2021 года ADNOC заключила контракт с инженерно-технологической компанией Technip Energies на разработку неиспользованных запасов нефти и газа на концессионных месторождениях Гаша, включая CCS в разработку.
Проекты и разработки УХУ – влияние иностранных доноров
Во многих регионах Азии первые проекты УХУ были запущены иностранными донорами. Не все эти проекты сохранились, но они, безусловно, повлияли на развитие CCS в Азии.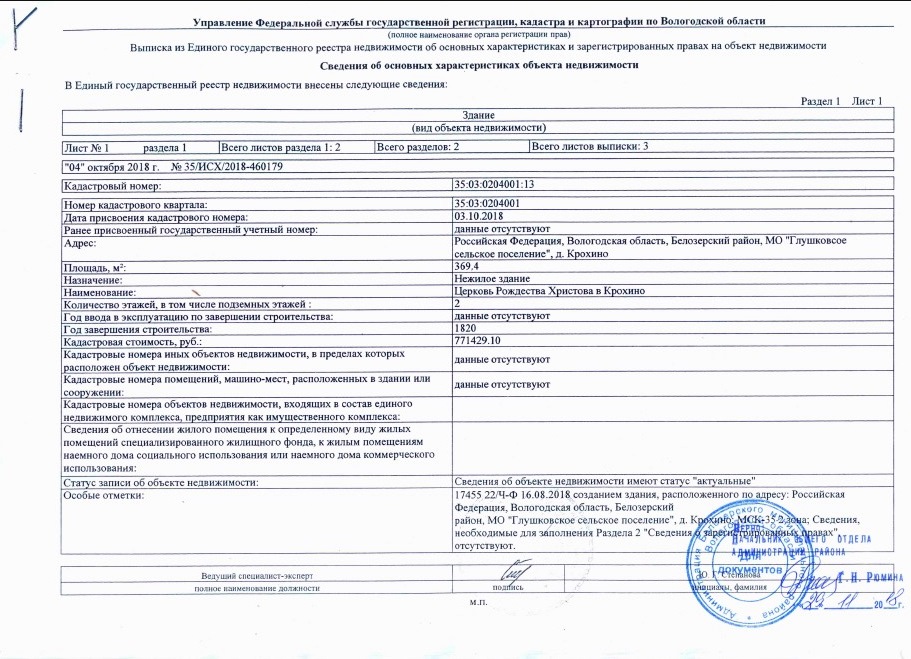 В настоящее время Индонезия является примером того, как индустрия УХУ начала развиваться благодаря внешним донорам. В других странах, особенно в Китае, этот этап давно завершился, и промышленность на местах создала импульс. Большинство спонсоров, государственных и частных, поступают из Австралии, Европейского Союза и Великобритании.
В настоящее время Индонезия является примером того, как индустрия УХУ начала развиваться благодаря внешним донорам. В других странах, особенно в Китае, этот этап давно завершился, и промышленность на местах создала импульс. Большинство спонсоров, государственных и частных, поступают из Австралии, Европейского Союза и Великобритании.
Китай
В 2010 году правительства Китая и Австралии согласовали технико-экономическое обоснование коммерческого развертывания CCS в Китае. Технология улавливания CO 2 на основе амина должна была быть испытана на двух электростанциях Huaneng-Group. Планы были поддержаны правительством Австралии в размере 11,5 млн австралийских долларов. Угольная электростанция Гаобэйдянь компании Huaneng недалеко от Пекина какое-то время рассматривалась как объект, но в конечном итоге запланированные проекты захвата не были реализованы.
В 2013 году Глобальный институт улавливания и хранения углерода поддержал проект Yanchang Petroleum CCS на сумму 2,5 миллиона австралийских долларов. Проект управляется государственной компанией Yanchang Petroleum и расположен в городе Юйлинь в провинции Шэньси. С 2012 года в рамках проекта работает небольшая установка по улавливанию CO 2 , в 2014 году было объявлено о крупномасштабном проекте, а в 2017 году началось ее строительство. Ввод в эксплуатацию более крупной установки был объявлен на 2018 год и с тех пор откладывается. Yanchang Petroleum планирует использовать захваченный CO 2 для увеличения нефтеотдачи в бассейне Ордос в центральном Китае.
Проект управляется государственной компанией Yanchang Petroleum и расположен в городе Юйлинь в провинции Шэньси. С 2012 года в рамках проекта работает небольшая установка по улавливанию CO 2 , в 2014 году было объявлено о крупномасштабном проекте, а в 2017 году началось ее строительство. Ввод в эксплуатацию более крупной установки был объявлен на 2018 год и с тех пор откладывается. Yanchang Petroleum планирует использовать захваченный CO 2 для увеличения нефтеотдачи в бассейне Ордос в центральном Китае.
С 2017 года ЕС финансирует проект «Китайско-европейские решения по сокращению выбросов» (CHEERS) на сумму 9,7 млн евро. Проект возглавляет норвежская компания SINTEF Energy AS и осуществляется в сотрудничестве с партнерами из Бельгии, Китая, Франции, Норвегии и Польши. CHEERS нацелен на тестирование, демонстрацию и дальнейшее развитие новых технологий улавливания CO 2 в нефтеперерабатывающей и других энергоемких отраслях. Среди партнеров проекта — научно-исследовательские институты и партнеры по ископаемой промышленности, такие как TOTAL и DONGFANG Boiler Group.
В рамках проекта Net Zero Emission Coal была предложена угольная электростанция с улавливанием CO 2 рядом с нефтяными месторождениями Дацин, Цзилинь или Цзянсу. Этот проект был совместным проектом Китая, ЕС, Великобритании и Норвегии и был поддержан государственным финансированием в размере более 16 миллионов долларов США из Норвегии и Великобритании. В 2007 году началось технико-экономическое обоснование, и угольная электростанция с улавливанием CO 2 должна была быть введена в эксплуатацию в 2014 году, но планы в рамках проекта не были реализованы.
Британско-китайский центр CCUS в Гуандуне (GDCCUSC), финансируемый государством Великобританией и Китаем, был создан в 2013 году для тестирования и продвижения CCS и CCUS в Китае. В состав центра входят исследовательские институты и компании, работающие на ископаемом топливе, в том числе Китайская национальная морская нефтяная корпорация, Alstom и Shell Cansolv. Центр реализует проекты CCS/CCUS на электростанции в Хайфэне и нефтеперерабатывающем заводе в Хуэйчжоу. В апреле 2022 года сотрудники GDCCUSC провели предварительное геологическое исследование для запланированной демонстрации минерализации CO 9.0265 2 в базальтовых породах на полуострове Чжаньцзян Лэйчжоу. Центр растет — в июне 2022 года GDCCUSC опубликовал несколько объявлений о вакансиях, в том числе для исследователей и инженеров в области геологического хранения и использования CO 2 .
В апреле 2022 года сотрудники GDCCUSC провели предварительное геологическое исследование для запланированной демонстрации минерализации CO 9.0265 2 в базальтовых породах на полуострове Чжаньцзян Лэйчжоу. Центр растет — в июне 2022 года GDCCUSC опубликовал несколько объявлений о вакансиях, в том числе для исследователей и инженеров в области геологического хранения и использования CO 2 .
Нефтегазовая климатическая инициатива (OGCI) направлена на поддержку развития пяти центров CCS в Китае, включая центр Xinjiang Jungger. Хаб Xinjiang Jungger должен был улавливать CO 2 с цементных, химических и электростанций. Китайская национальная нефтяная корпорация (CNPC) планировала инвестировать в проект CO 2 инфраструктура, и OGCI планировали поддерживать закачку CO 2 и деятельность по повышению нефтеотдачи. Партнеры по проекту также планировали работать с соответствующими министерствами Китая над созданием правовой базы для центров CCS. Точное местонахождение центра Xinjiang Jungger не разглашается. Весь проект был заброшен примерно в 2022 году, причина пока не разглашается.
Точное местонахождение центра Xinjiang Jungger не разглашается. Весь проект был заброшен примерно в 2022 году, причина пока не разглашается.
Индия
Министерство энергетики и изменения климата Великобритании профинансировало испытание технологии улавливания CO 2 на химическом заводе Solvay Chemicals в Индии Solvay Vishnu недалеко от Тирупати на сумму около 3,9 евро.млн. В качестве технологии использовалась технология химического растворителя от Carbon Clean, британского разработчика технологии улавливания CO 2 . Испытания начались в 2012 году, но установка по улавливанию CO 2 больше не входит в портфолио Solvay Chemicals.
Индонезия
Проект Gundih CCS финансируется посольством Норвегии, Киотским университетом и другими государственными учреждениями. В течение этого десятилетия проект направлен на проведение CO 2 инъекций в районе Гунди в провинции Центральная Ява для изучения CCS.
В 2020 году испанская нефтегазовая компания Repsol SA намеревалась провести исследование для крупномасштабного проекта CCS/CCUS в своих запасах природного газа на блоке Сакакеманг на Южной Суматре. В рамках проекта планируется закачка уловленного CO 2 на соседние месторождения Дайунг и Гелам.
В 2021 году Японская национальная корпорация нефти, газа и металлов, корпорация Mitsubishi и другие учреждения подписали меморандум о взаимопонимании и согласились провести совместное исследование CCS и CCUS на аммиачном заводе PAU в Лувуке, Центральный Сулавеси.
В июне 2021 года индонезийская государственная компания Pertamina подписала меморандум о взаимопонимании (МоВ) с Japan Petroleum Exploration Co. и расположенным в Джакарте центром исследований и разработок Lemigas для разработки CCS/CCUS на нефтегазовых месторождениях Суковати и Гундих. Pertamina подписала дополнительные меморандумы о взаимопонимании с ExxonMobil в ноябре 2021 года и с Air Liquide в январе 2022 года.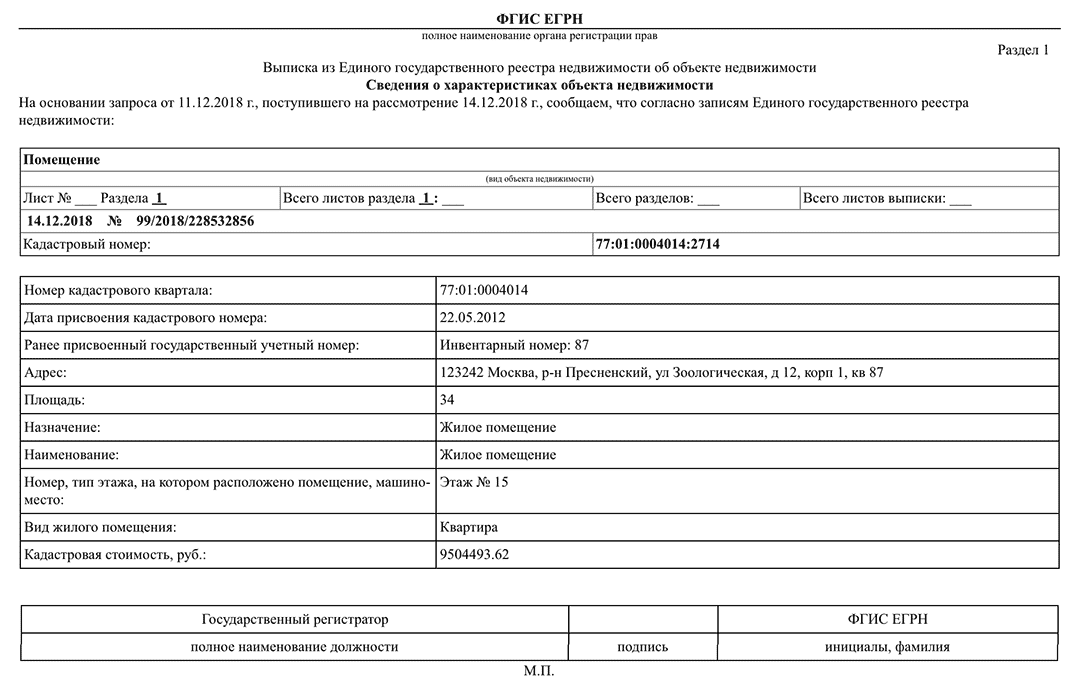
В 2021 году BP объявила, что изучит возможность использования CCS/CCUS на своем заводе по производству СПГ Tangguh в городе Телук Бинтуни, провинция Папуа-Барат. Проект включает СО 2 улавливание в Tangguh LNG, CO 2 транспортировка и повышенная добыча газа (EGR) на газовых месторождениях Убадари и Ворвата. Начиная с 2022 года в рамках проекта начнется этап начального проектирования и проектирования (FEED).
Израиль
ЕС профинансировал полевой эксперимент в рамках исследовательского проекта «МУСТАНГ — подход с множественными пространственными и временными масштабами для количественной оценки глубоких солевых образований для хранения CO 2 » с 2009 по 2014 год. Проект возглавлялся Шведским университетом Упсалы и проводился в сотрудничестве с 19партнеров по исследованиям из Европы и Израиля. Экспериментальный участок в Хелец, Израиль, использовался для инъекций CO 2 и для тестирования новых технологий мониторинга и измерения. Испытания проводились на окраинах истощенного нефтяного пласта на нефтяном месторождении Хелец в Израиле. Количество введенного CO 2 недоступно.
Испытания проводились на окраинах истощенного нефтяного пласта на нефтяном месторождении Хелец в Израиле. Количество введенного CO 2 недоступно.
Россия
В 2021 году российская нефтяная компания «Роснефть» подписала несколько соглашений о стратегическом сотрудничестве для развития совместных проектов CCS/CCUS, например, с BP, Shell и Equinor. В 2022 году из-за политической ситуации международные компании вышли из российских совместных предприятий.
Таиланд
В апреле 2022 года японская корпорация INPEX и японская корпорация JGC Holdings подписали меморандум о взаимопонимании с таиландской государственной компанией Petroleum Exploration and Production Public Company Ltd (PTTEP). Совместный проект направлен на изучение потенциальной разработки решений CCS для нефтегазового сектора и других отраслей с высоким потреблением ископаемого топлива.
CCUS – текущие разработки в азиатском контексте
Количество проектов по использованию и хранению углерода (CCUS) в Азии медленно, но неуклонно растет. Увеличивается собственная разработка технологии CCUS, но на большинстве площадок технология импортируется с других континентов, например CO 2 Технология улавливания от британской компании Carbon Clean. Проекты CCUS реализуются в химической промышленности, в пищевой промышленности, на металлургических и цементных заводах.
Увеличивается собственная разработка технологии CCUS, но на большинстве площадок технология импортируется с других континентов, например CO 2 Технология улавливания от британской компании Carbon Clean. Проекты CCUS реализуются в химической промышленности, в пищевой промышленности, на металлургических и цементных заводах.
Китай
Китайско-канадская компания C4X (China Canada CO 2 Conversion X) Technologies Inc. участвовала в конкурсе COSIA Carbon XPRIZE и вышла на финальный этап в 2019 году. электростанции, горнодобывающая промышленность, производство цемента, производство стали, винодельня и т. д. в химикаты и пластмассы, такие как этиленкарбонат, этиленгликоль, метанол и т. д.
В 2021 году исландская компания Carbon Recycling International начала установку своей системы производства метанола CO 2 , реакторной системы с выбросами в жидкость (ETL), рядом с предприятием по производству коксового газа в городе Аньянг. в провинции Хэнань. Завод будет эксплуатироваться проектной компанией Shunli и должен быть введен в эксплуатацию в третьем квартале 2022 года. Еще одна система ETL-реакторов планируется для нефтехимического комплекса в Ляньюньгане в провинции Цзянсу на восточном побережье Китая. СО 2 будет улавливаться на заводе по производству окиси этилена. Завод по производству метанола CO 2 будет принадлежать и эксплуатироваться Jiangsu Sailboat, и его ввод в эксплуатацию запланирован на 2023 год.
в провинции Хэнань. Завод будет эксплуатироваться проектной компанией Shunli и должен быть введен в эксплуатацию в третьем квартале 2022 года. Еще одна система ETL-реакторов планируется для нефтехимического комплекса в Ляньюньгане в провинции Цзянсу на восточном побережье Китая. СО 2 будет улавливаться на заводе по производству окиси этилена. Завод по производству метанола CO 2 будет принадлежать и эксплуатироваться Jiangsu Sailboat, и его ввод в эксплуатацию запланирован на 2023 год.
Индия
преобразовать этанол в топливо и омега-3 жирные кислоты в сотрудничестве с американской компанией LanzaTech. Планировалось строительство завода в промышленных масштабах, но нет никаких признаков того, что этот план все еще реализуется.
В 2019 году индийский производитель цемента Dalmia Cement Ltd объявил о подписании меморандума о взаимопонимании с британской компанией Carbon Clean. Dalmia Cement и Carbon Clean намерены построить установку по улавливанию CO 2 на заводе Dalmia Cement в штате Тамил Наду. Захваченный CO 2 будет использоваться для приложений CCUS или продан. Предложение было рассмотрено Азиатским банком развития в 2021 году, и Dalmia Cement недавно объявила о планах обратиться за финансовой помощью к правительству США.
Захваченный CO 2 будет использоваться для приложений CCUS или продан. Предложение было рассмотрено Азиатским банком развития в 2021 году, и Dalmia Cement недавно объявила о планах обратиться за финансовой помощью к правительству США.
Компания Breathe, основанная в 2016 году и базирующаяся в Бангалоре, перерабатывает захваченный CO 2 на метанол высокой чистоты и монооксид углерода. Компания Breathe приняла участие в конкурсе COSIA Carbon XPRIZE и вышла в финальный этап конкурса в 2019 году. Электростанция Виндхьячал, крупнейшая тепловая электростанция Индии. Технология улавливания CO 2 будет предоставлена British Carbon Clean, а компания Green Power International заключила контракт на проектирование и строительство CO 9.0265 2 завод по захвату. NETRA, научно-исследовательское подразделение NTPC, планирует создать на том же объекте демонстрационный завод по преобразованию CO 2 в метанол.
Tata Steel ввела в эксплуатацию небольшой проект по улавливанию CO 2 на своем сталелитейном заводе в Джамшедпуре в 2021 году. Технология улавливания CO 2 была предоставлена компанией Carbon Clean. Tata Steel планирует повторно использовать захваченный CO 2 на месте.
Технология улавливания CO 2 была предоставлена компанией Carbon Clean. Tata Steel планирует повторно использовать захваченный CO 2 на месте.
Япония
С 2019 г., Министерство окружающей среды Японии финансирует демонстрационный проект захвата CO 2 на заводе Taiheiyo Cement в Кумагая в городе Кумагая, префектура Сайтама. Демонстрационные испытания улавливания CO 2 на основе аминов начались осенью 2021 года. Для улавливания CO 2 Taiheiyo Cement выбрала технологию улавливания, разработанную Carbon Clean. В 2021 году компания Taiheiyo Cement получила грант в рамках проекта «Разработка технологии циркуляции углерода для цементной промышленности», который финансируется Японской организацией по развитию новых энергетических и промышленных технологий (NEDO).
Японская компания Mitsui Chemicals построила демонстрационную установку по производству метанола из промышленных стоков CO 2 . Завод расположен на объектах компании в Осаке, где ежегодно может улавливаться ~ 150 тонн CO 2 . Компания Mitsui запустила этот проект в сотрудничестве с Научно-исследовательским институтом инновационных технологий Земли в Киото. В 2021 году Mitsui Chemicals и Университет Кюсю совместно создали Центр углеродно-нейтральных исследований с целью исследования зеленого водорода, CO 2 — захват, а CO 2 — использование технологий.
Завод расположен на объектах компании в Осаке, где ежегодно может улавливаться ~ 150 тонн CO 2 . Компания Mitsui запустила этот проект в сотрудничестве с Научно-исследовательским институтом инновационных технологий Земли в Киото. В 2021 году Mitsui Chemicals и Университет Кюсю совместно создали Центр углеродно-нейтральных исследований с целью исследования зеленого водорода, CO 2 — захват, а CO 2 — использование технологий.
В апреле 2022 года компании Sekisui Chemical и LanzaTech Inc. завершили строительство демонстрационной установки по улавливанию CO 2 на заводе Sekisui Chemical по производству этанола в городе Кудзи, префектура Иватэ. LanzaTech разработала технологию CCUS, которая улавливает CO 2 из выхлопных газов и преобразует его в этанол. Преобразование осуществляется микробами, которые питаются CO 2 и производят этанол. Sekisui Chemical стремится проверить техническую и экономическую осуществимость технологии. Демонстрационная установка составляет примерно одну десятую от размера будущей коммерческой установки. С 2021 года Sekisui Chemical также работает с ArcelorMittal над проектом по улавливанию и повторному использованию CO 9.0265 2 от производства стали.
Демонстрационная установка составляет примерно одну десятую от размера будущей коммерческой установки. С 2021 года Sekisui Chemical также работает с ArcelorMittal над проектом по улавливанию и повторному использованию CO 9.0265 2 от производства стали.
Саудовская Аравия
В 2021 году Gulf Cryo и итальянская Tecno Project Industriale подписали меморандум о взаимопонимании по строительству и эксплуатации завода по улавливанию CO 2 с целью удвоить существующие производственные мощности Gulf Cryo по CO 2 в Саудовской Аравии. Аравия. Ожидается, что завод будет улавливать ~ 0,25 миллиона тонн CO 2 в год для производства высокочистого пищевого CO 2 . Gulf Cryo — производитель промышленных газов, в том числе сжатого CO 9, в Объединенных Арабских Эмиратах.0265 2 в газообразной или жидкой форме, различной концентрации и чистоты. CO 2 , вероятно, будет перехвачен на заводе Gulf Cryo в Эр-Рияде.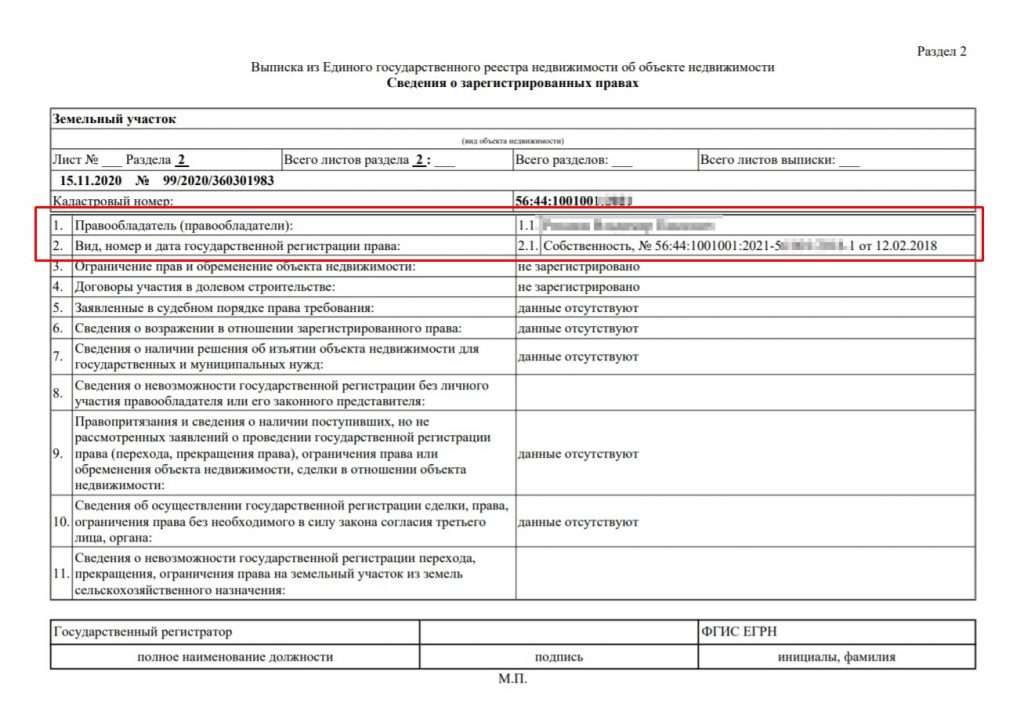 Пока нет информации об используемой технологии улавливания CO 2 или точном графике. У Gulf Cryo есть дополнительные проекты по улавливанию CO 2 в Кувейте и Саудовской Аравии.
Пока нет информации об используемой технологии улавливания CO 2 или точном графике. У Gulf Cryo есть дополнительные проекты по улавливанию CO 2 в Кувейте и Саудовской Аравии.
Компания Sustainable Energy Solutions (США) и Исследовательский центр чистого сгорания при Университете науки и технологий имени короля Абдуллы (KAUST) (Саудовская Аравия) разработали и продемонстрировали технологию криогенного улавливания углерода (CCC). CCC улавливает CO 2 от выхлопных газов в процессе дожигания. При этом выхлопные газы охлаждаются до температуры ниже -130°C. Таким образом, CO 2 замораживается, затвердевает и отделяется от выхлопных газов. Затем CO 2 сжижается и подготавливается к транспортировке. В 2021 году было завершено предварительное исследование по развертыванию системы CCC в Исследовательском центре чистого сгорания KAUST в Дубае, Саудовская Аравия. Ожидается, что планируемая установка будет улавливать до 0,011 млн тонн CO 9.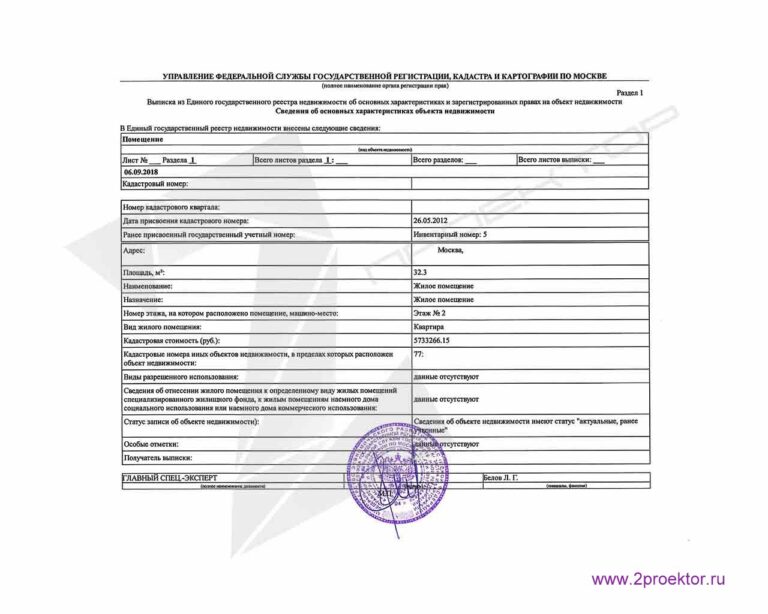 0265 2 ежегодно. По данным KAUST, Национальная программа экономики замкнутого цикла углерода в Саудовской Аравии, инициатива Министерства энергетики, активно занимается коммерциализацией технологии CCC в Саудовской Аравии.
0265 2 ежегодно. По данным KAUST, Национальная программа экономики замкнутого цикла углерода в Саудовской Аравии, инициатива Министерства энергетики, активно занимается коммерциализацией технологии CCC в Саудовской Аравии.
Тайвань
В 2014 году компания LanzaTech в сотрудничестве с White BioTech (WBT) открыла демонстрационный промышленный завод по преобразованию выхлопных газов стали в этанол в Гаосюне, Тайвань. Демонстрационная установка производила около 100 килограммов этанола в сутки. Планы коммерческого завода были окончательно согласованы в апреле 2015 года, но нет никаких признаков того, что они были реализованы.
Объединенные Арабские Эмираты
В марте 2022 года правительство Германии подписало соглашения о сотрудничестве с Объединенными Арабскими Эмиратами по производству голубого водорода, который должен быть доставлен в Германию уже в 2022 году.
Государственная нефтяная компания Mubadala Petroleum и итальянская Eni объявили о соглашении о совместном изучении CCUS в сентябре 2021 года.
В декабре 2021 года Национальная нефтяная компания Абу-Даби подписала соглашение с французской TotalEnergies о совместном изучении CCS/CCUS.
Многие проекты CCUS все еще находятся в разработке или на ранних стадиях. Несколько текущих проектов показывают, что некоторые отрасли заинтересованы в CO 2 в качестве сырья в ограниченном количестве. Однако это не означает, что продукты, в которых используется CO 2 , подходят для хранения CO 2 . Улавливание, транспортировка и очистка уловленного CO 2 очень энергоемки, и уловленный CO 2 выбрасывается обратно в атмосферу, как только CO 2 — содержащие продукты, такие как химикаты или газированная вода. Кроме того, не следует забывать, что CO 2 производится заранее – из ископаемого топлива, которое, помимо вредных для климата газов, оказывает много других негативных воздействий на окружающую среду и здоровье.
Индукция семейства факторов транскрипции Early Growth Response (Egr) при селекции тимуса | Журнал экспериментальной медицины
Skip Nav Destination
Статья|
17 февраля 1997 г.
Хуэй Шао,
Дуайт Х. Коно,
Линг-Ю Чен,
Элисса М. Рубин,
Джонатан Кэй
Информация об авторе и статье
Адресная переписка с доктором Джонатаном Кеем, отделение иммунологии, IMM-8, Научно-исследовательский институт Скриппса, 10550 N. Torrey Pines Rd., La Jolla, California
.
1 Сокращения, использованные в данной статье: CsA, циклоспорин А; Erg-1, реакция раннего роста.
Х. Шао и Д. Коно внесли равный вклад в эту работу.
Полученный:
10 октября 1996 г.
Полученная редакция:
09 декабря 1996 г.
Номер для печати в Интернете: 1540-9538
Номер для печати: 0022-1007
J Exp Med (1997) 185 (4): 731–744.
https://doi.org/10.1084/jem.185.4.731
История статьи
Получено:
10 октября 1996 г.
Пересмотр получено:
09 декабря 1996 г.
- Стандартный вид
Просмотры
- Содержание артикула
- Рисунки и таблицы
- Видео
- Аудио
- Дополнительные данные
- Экспертная оценка
PDF
Делиться
- MailTo
- Твиттер
Инструменты
Получить разрешения
Иконка Цитировать
Цитировать
Поиск по сайту
Citation
Хуэй Шао, Дуайт Х. Коно, Линг-Ю Чен, Элисса М. Рубин, Джонатан Кэй; Индукция семейства факторов транскрипции Early Growth Response (Egr) во время тимической селекции. J Exp Med 17 февраля 1997 г.; 185 (4): 731–744. doi: https://doi.org/10.1084/jem.185.4.731
Коно, Линг-Ю Чен, Элисса М. Рубин, Джонатан Кэй; Индукция семейства факторов транскрипции Early Growth Response (Egr) во время тимической селекции. J Exp Med 17 февраля 1997 г.; 185 (4): 731–744. doi: https://doi.org/10.1084/jem.185.4.731
Скачать файл цитаты:
- Рис (Зотеро)
- Диспетчер ссылок
- EasyBib
- Подставки для книг
- Менделей
- Бумаги
- Конечная примечание
- RefWorks
- Бибтекс
панель инструментов поиска
Расширенный поиск
Мало что известно о регуляции экспрессии генов во время TCR-опосредованной дифференцировки незрелых CD4 + 8 + (двойной положительный) тимоцитов в зрелые Т-клетки. Используя линию клеток-предшественников тимоцитов DPK CD4 + 8 + , мы демонстрируем, что ген раннего ответа роста-1 ( Erg-1 ), кодирующий фактор транскрипции цинкового пальца, быстро активируется после стимуляции TCR. Мы также сообщаем, что Egr-1 экспрессируется подмножеством нормальных двойных положительных тимоцитов в коре тимуса, а также большинством медуллярных одиночных положительных тимоцитов. Экспрессия Egr-1 резко снижена в тимусе мышей с нокаутом главного комплекса гистосовместимости, но может быть индуцирована стимуляцией анти-CD3-антителами изолированных тимоцитов этих животных. Эти и другие данные свидетельствуют о том, что высокий уровень экспрессии Egr-1 в тимусе является следствием отбора. Сходный паттерн экспрессии обнаружен у членов семейства Egr-2 и Egr-3. Используя клеточную линию DPK, мы также демонстрируем, что экспрессия Egr-1, 2 и 3 зависит от активации ras, как и инициация дифференцировки в одну положительную клетку. Напротив, ингибитор кальциневрина циклоспорин А, который ингибирует дифференцировку клеток DPK, а также положительный отбор, ингибирует экспрессию Egr-2 и Egr-3, но не Egr-1. Идентификация семейства Egr в этом контексте представляет собой первое сообщение о связи между двумя известными сигнальными путями, участвующими в положительном отборе и нижестоящими регуляторами транскрипции.
Мы также сообщаем, что Egr-1 экспрессируется подмножеством нормальных двойных положительных тимоцитов в коре тимуса, а также большинством медуллярных одиночных положительных тимоцитов. Экспрессия Egr-1 резко снижена в тимусе мышей с нокаутом главного комплекса гистосовместимости, но может быть индуцирована стимуляцией анти-CD3-антителами изолированных тимоцитов этих животных. Эти и другие данные свидетельствуют о том, что высокий уровень экспрессии Egr-1 в тимусе является следствием отбора. Сходный паттерн экспрессии обнаружен у членов семейства Egr-2 и Egr-3. Используя клеточную линию DPK, мы также демонстрируем, что экспрессия Egr-1, 2 и 3 зависит от активации ras, как и инициация дифференцировки в одну положительную клетку. Напротив, ингибитор кальциневрина циклоспорин А, который ингибирует дифференцировку клеток DPK, а также положительный отбор, ингибирует экспрессию Egr-2 и Egr-3, но не Egr-1. Идентификация семейства Egr в этом контексте представляет собой первое сообщение о связи между двумя известными сигнальными путями, участвующими в положительном отборе и нижестоящими регуляторами транскрипции.
Развитие незрелых CD4 + 8 + (двойных положительных) тимоцитов в зрелые CD4 + или CD8 + (одиночные положительные) Т-клетки инициируется TCR-опосредованным распознаванием молекул MHC, экспрессируемых тимусом. строма. Хотя этот процесс, называемый положительным отбором, сопровождается рядом легко наблюдаемых изменений в экспрессии белков клеточной поверхности, в первую очередь потерей корецептора CD4 или CD8, мало что известно о контроле этого события дифференцировки, опосредованного рецептором, на уровне регуляции генов. Конечно, внутриклеточные сигналы, генерируемые TCR, должны быстро изменять активность и/или экспрессию регуляторов транскрипции, которые затем вызывают каскад специфических для типа клеток изменений в экспрессии генов, необходимых для установления фенотипа зрелых Т-клеток. Немедленные ранние гены, определяемые как те гены, экспрессия которых не зависит от синтеза белка de novo, являются вероятными кандидатами в качестве медиаторов быстрого ответа между сигналами, генерируемыми рецепторами клеточной поверхности, и последующими изменениями в экспрессии генов.
Реакция на ранний рост-1 ( Egr-1 1 ; также известная как zif268, Krox-24, tis-8, NGFI-A и pAT 225) является одним из таких генов немедленного раннего роста, который был независимо выделен из фибробластов. активируется сывороткой или форболовым эфиром (1–5), из клеток PC12, которые были индуцированы для дифференцировки нейронов с помощью обработки фактором роста нервов (6), и из активированных митогеном и форболовым эфиром Т-лимфоцитов периферической крови (7). Белок Egr-1 представляет собой регулятор транскрипции, содержащий три структурных домена цинковых пальцев Cys 9.0265 2 Тип His 2 (8), где каждый цинковый палец взаимодействует с двумя гуанинами в субсайте из трех пар оснований минимальной последовательности связывания нонамера (9–11). В дополнение к ДНК-связывающему домену в Egr-1 идентифицированы как активационные, так и ингибирующие области (12-14), последняя из которых связывает недавно идентифицированный белок-репрессор (15). Основываясь на сохранении ДНК-связывающих доменов, Egr-1 принадлежит к семейству факторов транскрипции, которое также включает Krox-20/Egr-2/pAT 591 (16–19), Egr-3 (20) и Egr-4 /NGFI-C/pAT133 (20–22). Члены семейства Egr-1 имеют около 90% идентичности аминокислот в домене цинковых пальцев.
Основываясь на сохранении ДНК-связывающих доменов, Egr-1 принадлежит к семейству факторов транскрипции, которое также включает Krox-20/Egr-2/pAT 591 (16–19), Egr-3 (20) и Egr-4 /NGFI-C/pAT133 (20–22). Члены семейства Egr-1 имеют около 90% идентичности аминокислот в домене цинковых пальцев.
Egr-1 экспрессируется различными типами клеток в ответ на митогены, сигналы дифференцировки, тканевое или радиационное повреждение или возбуждение нейронов (23). Широко распространенная экспрессия Egr-1 предполагает, что этот регулятор транскрипции может играть роль в связывании общих биохимических сигнальных путей с быстрыми изменениями экспрессии генов. Хотя функция Egr-1 еще предстоит выяснить в большинстве систем, было показано, что экспрессия Egr-1 в культивируемых гемопоэтических линиях клеток-предшественников блокирует дифференцировку гранулоцитов, хотя и позволяет дифференцироваться в линию макрофагов (24, 25), демонстрируя потенциал для этот фактор транскрипции воздействует на приверженность клеточной линии. Недавно также сообщалось, что Egr-1 регулирует группу генов, участвующих в факторе роста и реакции свертывания крови на повреждение эндотелиальных клеток (26, 27).
Недавно также сообщалось, что Egr-1 регулирует группу генов, участвующих в факторе роста и реакции свертывания крови на повреждение эндотелиальных клеток (26, 27).
В лимфоцитах Egr-1 и другие члены семейства индуцируются как в В-, так и в Т-клетках при перекрестном связывании антигенных рецепторов или в ответ на митогены или сложные эфиры форбола (1, 28–32). Интересно, что в В-лимфоцитах индуцируемая экспрессия Egr-1 коррелирует со стадией развития клетки. Таким образом, зрелые, но не незрелые В-клетки можно индуцировать для экспрессии Egr-1 при стимуляции эфирами форбола или антителами против IgM (33). Кроме того, перекрестное связывание Fc-рецепторов, которое может отрицательно регулировать опосредованную антииммуноглобулинами активацию В-клеток, ингибирует индукцию Egr-1 (34, 35). Вместе эти результаты указывают на регулируемую в процессе развития положительную роль Egr-1 в активации В-клеток. Однако IL-4 может преодолевать опосредованное рецептором Fc ингибирование пролиферации В-клеток без увеличения уровня мРНК Egr-1 (34).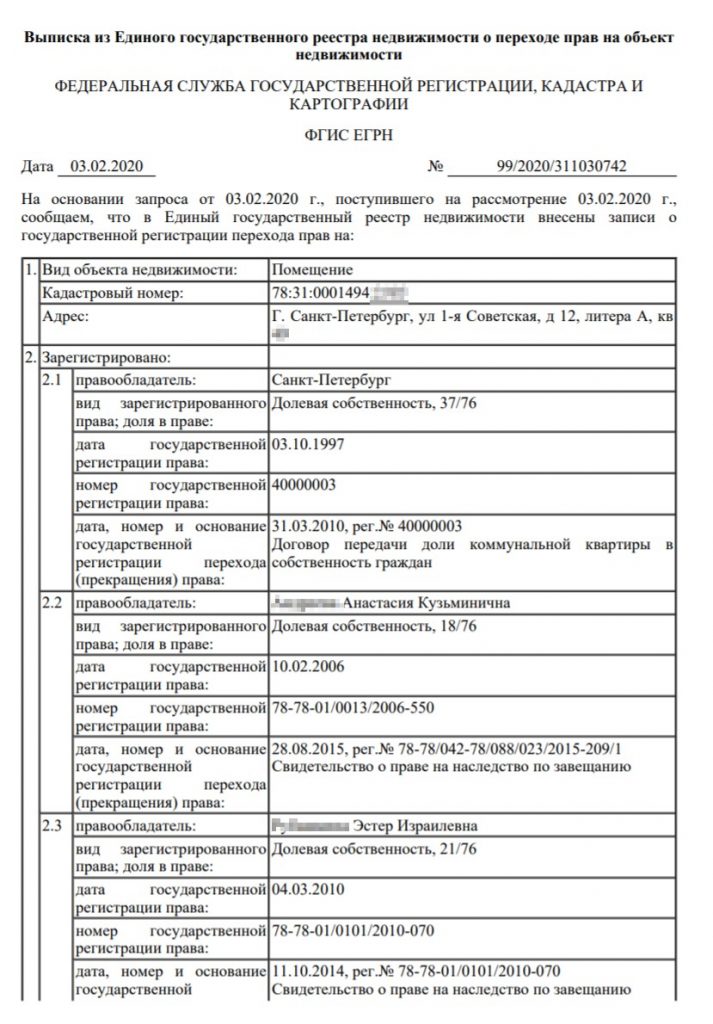 Таким образом, роль Egr-1 в функции зрелых лимфоцитов может зависеть от специфических средств клеточной активации. У взрослых мышей высокие уровни мРНК Egr-1 обнаруживаются в головном мозге, тимусе, сердечной мышце и легких, а также более низкие уровни в других тканях (2–4, 36, 37). Источник экспрессии тимуса Egr-1 ранее не исследовался.
Таким образом, роль Egr-1 в функции зрелых лимфоцитов может зависеть от специфических средств клеточной активации. У взрослых мышей высокие уровни мРНК Egr-1 обнаруживаются в головном мозге, тимусе, сердечной мышце и легких, а также более низкие уровни в других тканях (2–4, 36, 37). Источник экспрессии тимуса Egr-1 ранее не исследовался.
Мы описали систему культивирования in vitro с использованием клеточной линии DPK, которая позволяет нам анализировать опосредованный TCR переход двойной положительной Т-клетки в одиночную положительную CD4 Т-клетку (38). Используя метод субтрактивной гибридизации на основе ПЦР, мы идентифицировали Egr-1 как ген, который быстро индуцируется во время дифференцировки клеток DPK. Основываясь на этом открытии, мы проанализировали экспрессию Egr-1 в нормальном тимусе. Мы сообщаем здесь, что высокий уровень активности связывания мРНК, белка и ДНК Egr-1 связан с отбором тимуса и может представлять собой один из самых ранних маркеров вовлечения TCR в тимусе.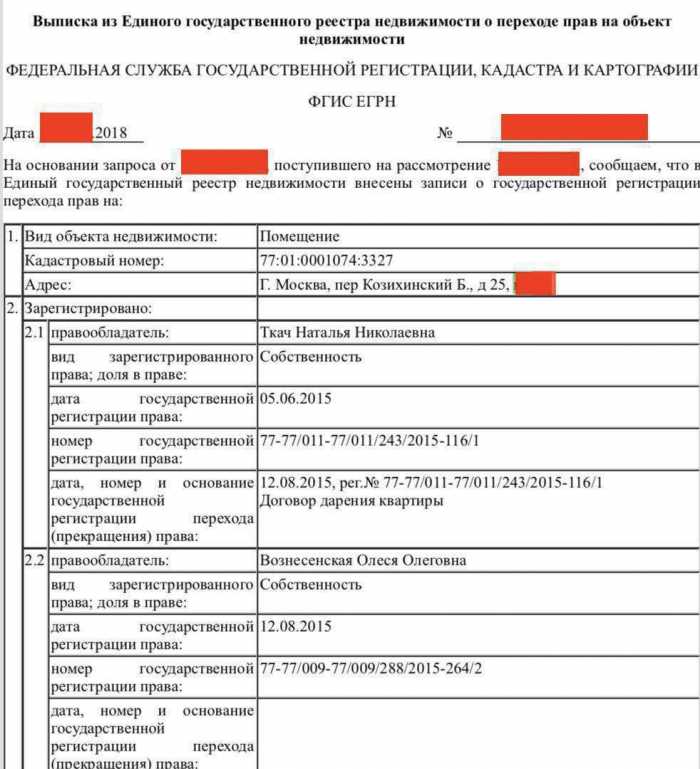 Мы также представляем доказательства того, что экспрессия семейства Egr в этом типе клеток зависит от путей передачи сигналов ras и calcineurin, как и дифференцировка незрелых Т-клеток. Наша идентификация семейства Egr в этом контексте представляет собой важную связь между сигнальными путями, участвующими в положительном отборе, и нижележащими регуляторами экспрессии генов.
Мы также представляем доказательства того, что экспрессия семейства Egr в этом типе клеток зависит от путей передачи сигналов ras и calcineurin, как и дифференцировка незрелых Т-клеток. Наша идентификация семейства Egr в этом контексте представляет собой важную связь между сигнальными путями, участвующими в положительном отборе, и нижележащими регуляторами экспрессии генов.
Мыши.
Мыши были получены из колонии грызунов Научно-исследовательского института Скриппса. Мыши с дефицитом MHC были получены путем скрещивания мышей с нокаутом по MHC класса II (39) с мышами с нокаутом по микроглобулину β2 (40) и были получены от доктора Дж. Спрента (Исследовательский институт Скриппса).
Клетки, анализы и антигены.
Получение, поддержание и дифференцировка клеточной линии DPK были описаны ранее (38). Клеточные линии DPK, экспрессирующие H-rasN17, доминантно-негативный мутант p21ras, были созданы с помощью переноса генов, опосредованного ретровирусами, как описано ранее (41), с использованием конструкции pZip-RasH(17N), щедро предоставленной доктором С. Дером (Университет Северной Каролины). в Чапел-Хилле) (42). Вкратце, клетки DPK инфицировали совместным культивированием с ретровирусной упаковочной клеточной линией PA317, которая ранее была трансфицирована pZip-RasH(17N). Через 2 дня клетки DPK переносили с упаковочной линии в полную среду, содержащую G418. Полученные устойчивые к G418 клеточные линии DPK анализировали фенотипически и функционально, как описано в тексте. Клетки DPK, которые экспрессировали RasH(17N), выращивали в условиях, идентичных DPK дикого типа, и не было отмечено никаких изменений скорости роста или морфологии. DCEK-ICAM представляет собой клеточную линию фибробластов, трансфицированную MHC E9 класса II.0940 k и гены ICAM-1 (43).
Дером (Университет Северной Каролины). в Чапел-Хилле) (42). Вкратце, клетки DPK инфицировали совместным культивированием с ретровирусной упаковочной клеточной линией PA317, которая ранее была трансфицирована pZip-RasH(17N). Через 2 дня клетки DPK переносили с упаковочной линии в полную среду, содержащую G418. Полученные устойчивые к G418 клеточные линии DPK анализировали фенотипически и функционально, как описано в тексте. Клетки DPK, которые экспрессировали RasH(17N), выращивали в условиях, идентичных DPK дикого типа, и не было отмечено никаких изменений скорости роста или морфологии. DCEK-ICAM представляет собой клеточную линию фибробластов, трансфицированную MHC E9 класса II.0940 k и гены ICAM-1 (43).
Клетки DPK активировали пептидом цитохрома с голубя 88104 (синтезирован в Научно-исследовательском институте Скриппса) и клетками DCEKICAM, как описано ранее (38). В некоторых экспериментах клетки DPK или тимоциты обрабатывали mAb 2C11 против CD3ε (PharMingen, Сан-Диего, Калифорния), предварительно адсорбированными на пластиковых чашках для культур тканей (10 мкг/мл) или 5-мкм полистироловыми латексными шариками (10 мкг 2C11 на 10 7 гранул) (Interfacial Dynamics Corporation, Портленд, Орегон).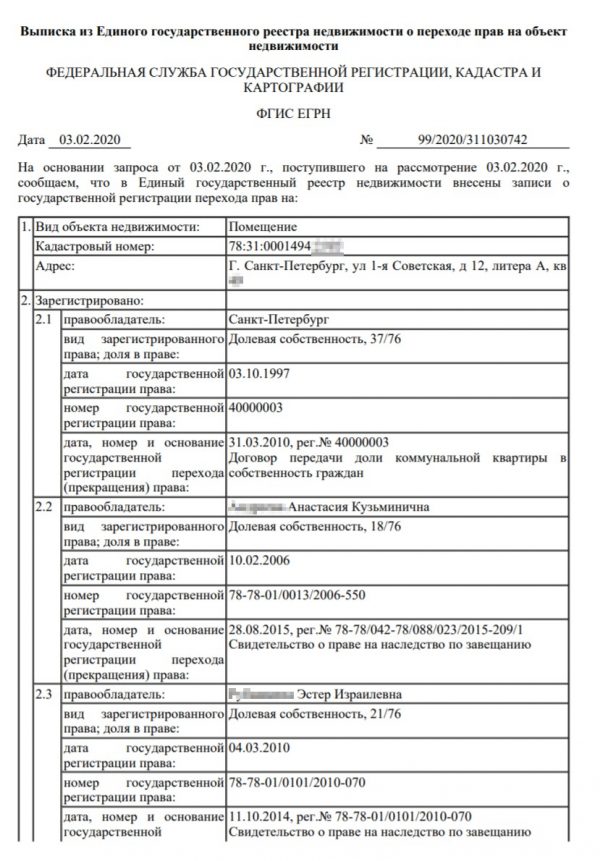 Для выделения РНК 1 × 10 9Клетки 0940 7 культивировали в 100-мм чашках для культивирования, покрытых 2C11, или с шариками, покрытыми 2C11, при соотношении клеток/гранул 1:1.
Для выделения РНК 1 × 10 9Клетки 0940 7 культивировали в 100-мм чашках для культивирования, покрытых 2C11, или с шариками, покрытыми 2C11, при соотношении клеток/гранул 1:1.
Вестерн-блот. Клетки
DPK лизировали в 1% NP-40 в PBS, содержащем 2 мкг/мл апротинина (Sigma Chem. Co., Сент-Луис, Миссури), 2 мкг/мл лейпептина (Sigma) и 1 мМ PMSF (Sigma). Клеточные лизаты инкубировали на льду в течение 15 минут, а постядерный супернатант извлекали микроцентрифугированием. 20 мкг общего белка подвергали SDS-PAGE (12%) и переносили на мембрану PVDF (Millipore Corp., Бедфорд, Массачусетс). Мембрану блокировали 5% обезжиренным молоком в промывочном буфере (0,15 М NaCl, 10 мМ Трис, pH 7,4, 2,5 мМ MgCl 9).0265 2 /0,5% Tween 20) и исследовали моноклональным антителом против Ras (Transduction Laboratories, Lexington, KY), разведенным в 2,5% нежирном молоке в промывочном буфере. Блот был тщательно промыт и проявлен с добавлением конъюгированного с пероксидазой козьего антимышиного IgG-антитела (Bio-Rad Laboratories, Hercules, CA) и визуализирован с использованием системы Enhanced Chemiluminescence Detection System (Amersham, Arlington Heights, IL).
Антитела и окрашивание.
Анти-CD4, конъюгированные с фикоэритрином (GIBCO BRL, Gaithersburg, MD), конъюгированные с Red613 (GIBCO BRL) или конъюгированные с цихромом (PharMingen) анти-CD8α, биотинилированные анти-CD69или mAb против CD3 (PharMingen) использовали в анализе FACS ® . Окрашивание клеточной поверхности выполняли, как описано ранее (38), и окрашенные клетки анализировали на FACScan ® или FACSort ® с использованием программного обеспечения CellQuest (Becton Dickinson, Mountain View, CA). Показана логарифмическая флуоресценция 5 000–20 000 жизнеспособных клеток, гейтированная в соответствии с их боковым и прямым светорассеянием.
Для четырехцветного анализа FACS ® тимоциты (3 × 10 6 ) молодой взрослой мыши B10.BR были окрашены по стандартной методике анти-CD4-PE и анти-CD8-цихромом в сочетании с биотинилированные анти-CD69или анти-CD3, а затем APCстрептавидин (Biomedia, Foster City, CA). Для окрашивания на Egr-1 клетки промывали в буфере для окрашивания (PBS, 3%, FCS, 0,1% азида) и фиксировали в 200 мкл 3% формальдегида в PBS в течение 30 мин при 4°C. Клетки отмывали в буфере для окрашивания, ресуспендировали в 100 мкл 0,5% тритона Х-100 в PBS и инкубировали 10 мин при комнатной температуре. После отмывки в окрашивающем буфере клетки ресуспендировали в 100 мкл блокирующего буфера (5% козья сыворотка в 0,1 М Трис, рН 7,2, 0,01% Тритон Х-100) и инкубировали еще 10 мин. Антисыворотка против пептида Egr-1 (C-19) (Santa Cruz Biotechnology, Санта-Крус, Калифорния) добавляли к клеткам в конечном разведении 1:1200 и продолжали инкубацию в течение дополнительных 30 минут при комнатной температуре. Затем клетки промывали и инкубировали с FITC-конъюгированным F(ab’) 2 против IgG кролика (Jackson ImmunoResearch Labs., West Grove, PA) в течение 30 мин при комнатной температуре. После окончательной промывки окрашенные клетки анализировали на FACSort ® , модернизированном до FACSCalibre ® с добавлением второго диодного лазера и детектора FL4.
Для окрашивания на Egr-1 клетки промывали в буфере для окрашивания (PBS, 3%, FCS, 0,1% азида) и фиксировали в 200 мкл 3% формальдегида в PBS в течение 30 мин при 4°C. Клетки отмывали в буфере для окрашивания, ресуспендировали в 100 мкл 0,5% тритона Х-100 в PBS и инкубировали 10 мин при комнатной температуре. После отмывки в окрашивающем буфере клетки ресуспендировали в 100 мкл блокирующего буфера (5% козья сыворотка в 0,1 М Трис, рН 7,2, 0,01% Тритон Х-100) и инкубировали еще 10 мин. Антисыворотка против пептида Egr-1 (C-19) (Santa Cruz Biotechnology, Санта-Крус, Калифорния) добавляли к клеткам в конечном разведении 1:1200 и продолжали инкубацию в течение дополнительных 30 минут при комнатной температуре. Затем клетки промывали и инкубировали с FITC-конъюгированным F(ab’) 2 против IgG кролика (Jackson ImmunoResearch Labs., West Grove, PA) в течение 30 мин при комнатной температуре. После окончательной промывки окрашенные клетки анализировали на FACSort ® , модернизированном до FACSCalibre ® с добавлением второго диодного лазера и детектора FL4.
Для иммуногистологии криостатные тонкие срезы (8–10 мк) тимуса фиксировали в 3% формальдегиде на PBS, инкубировали с 0,5% Тритоном Х-100 в течение 10 мин и блокировали в течение 10 мин в 5% козьей сыворотке в 0,1 М Трис, рН 7,2, содержащий 0,01% тритона Х-100. Срезы окрашивали специфической аффинно-очищенной кроличьей антисывороткой против пептида Egr-1 (Santa Cruz Biotechnology) в течение 1 часа, а затем пероксидазно-связанной антисывороткой F(ab’) 2 осла против кроличьего IgG (Jackson ImmunoResearch Labs). в течение 30 мин. Для контроля окрашивания антисыворотку к пептиду Egr-1 предварительно инкубировали в течение 2 ч при комнатной температуре с 10-кратным избытком по массе иммунизирующего пептида (Santa Cruz Biotechnology). Окрашивание осуществляли тетрагидрохлоридом 3,3′-диаминобензидина (Pierce, Рокфорд, Иллинойс), и срезы слегка контрастировали в растворе гематоксилина Гилла номер 2 (Sigma) с последующей обработкой кислым спиртом и бикарбонатом натрия. Срезы обезвоживали в спирте с возрастающей концентрацией, очищали Hemo-De (Fisher, Питтсбург, Пенсильвания) и заключали покровные стекла в монтажную среду DPX (BDH Laboratory Supplies, Poole, UK).
Репрезентативный анализ различий.
TRIzol Reagent (GIBCO BRL) использовали для получения суммарной РНК из клеток DPK через 6 ч после инкубации с клетками FcR + CHO (подарок доктора И. Меллмана, Йельский университет) в присутствии или в отсутствие растворимого анти-CD3ε 2C11. mAb. РНК Poly(A) + выделяли из тотальной РНК с использованием магнитных шариков oligo-dT (Dynal, Lake Success, NY). Двухцепочечную кДНК получали с использованием обратной транскриптазы SuperScript (GIBCO BRL) в соответствии с инструкциями производителя и расщепляли с помощью DpnII (New England Biolabs). Репрезентативный анализ различий был выполнен, как описано Hubank и Schatz (44). кДНК, полученная из клеток DPK, инкубированных с FcR 9В качестве драйвера использовали клетки СНО 0940 + , а в качестве тестера использовали кДНК, полученную из идентичных культур, но с добавлением mAb против CD3ε. Конечные продукты очищали из агарозных гелей, клонировали в клонирующий вектор ТА (Invitrogen, Сан-Диего, Калифорния) и секвенировали с помощью секвенирования цикла Taq (Perkin-Elmer, Норуолк, Коннектикут). Поиск сходства последовательностей идентифицировал четыре фрагмента 705, 671, 434 и 280 п.н. как фрагменты DpnII мышиного гена Egr-1 .
Поиск сходства последовательностей идентифицировал четыре фрагмента 705, 671, 434 и 280 п.н. как фрагменты DpnII мышиного гена Egr-1 .
ОТ-ПЦР.
Тотальную РНК получали из клеток DPK или тимоцитов с использованием реагента TRIzol (GIBCO BRL) или набора RNeasy RNA (Qiagen, Chatsworth, CA). Первую цепь кДНК, праймированную олиго-dT, получали из 1-4 мкг тотальной РНК с использованием системы предварительной амплификации SuperScript (GIBCO BRL) в соответствии с инструкциями производителя. ПЦР проводили в общем объеме 25 мкл, содержащем по 10 пмоль каждого праймера, 1× ПЦР-буфер (GIBCO BRL), 0,2 мМ dNTP, 2 мМ MgCl-9.0265 2 , 1 ЕД Taq-полимеразы (GIBCO BRL) и 1–5 мкл соответствующим образом разбавленной кДНК. Условия цикла: 94°С в течение 4 мин, затем 28–30 циклов 94°С в течение 30 с, 58 или 61°С (для Egr-3 ) в течение 40 с и 72°С в течение 1 мин. После заключительной инкубации при 72°С в течение 2 мин добавляли нагрузочный краситель и проводили реакции на 2,0% агарозном геле.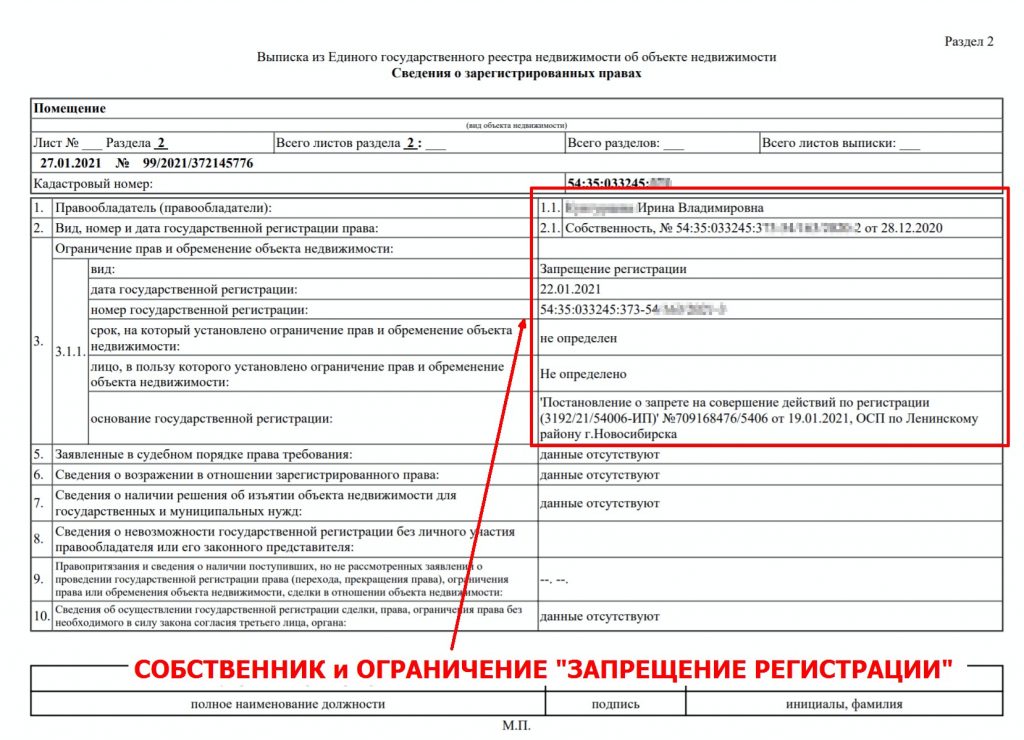 Продукты ПЦР визуализировали окрашиванием бромистым этидием.
Продукты ПЦР визуализировали окрашиванием бромистым этидием.
Конкурентную ОТ-ПЦР проводили, как описано выше, но с добавлением различных концентраций конкурентной конструкции Е4. E4 содержит оба Egr-1 и CD4 праймерные сайты, фланкирующие нерелевантную последовательность ДНК. Амплификация E4 с соответствующими праймерами приводит к продуктам ПЦР размером 395 п.н. (CD4) и 447 п.н. (Egr-1), в то время как кДНК CD4 и Egr-1 дает продукты ПЦР размером 486 п.н. и 356 п.н. соответственно. Для измерения концентрации конкретной кДНК в образце постоянное количество кДНК добавляли в реакционные пробирки, содержащие известные концентрации конкурента Е4. Отдельные реакции ПЦР проводили с праймерами CD4 и Egr-1. ПЦР-продукты конкурентов и кДНК разделяли электрофорезом в агарозном геле и окрашивали бромистым этидием. Флуоресцентные изображения были записаны на диск с помощью системы ImageStore 7500 (UVP, Inc., Upland, CA), а интенсивности полос были количественно оценены с использованием программного обеспечения NIH Image. Отношение конкурента к продукту ПЦР, полученному из кДНК, рассчитывали и наносили на график в зависимости от концентрации E4. Концентрацию специфической кДНК в образце принимали за концентрацию Е4, чтобы получить соотношение полос, равное 1. Для определения концентрации специфической кДНК в отдельном образце проводили как минимум два независимых конкурентных анализа ПЦР.
Отношение конкурента к продукту ПЦР, полученному из кДНК, рассчитывали и наносили на график в зависимости от концентрации E4. Концентрацию специфической кДНК в образце принимали за концентрацию Е4, чтобы получить соотношение полос, равное 1. Для определения концентрации специфической кДНК в отдельном образце проводили как минимум два независимых конкурентных анализа ПЦР.
Последовательности праймеров выше и ниже по течению, используемые в ОТ-ПЦР, были получены из разных экзонов. Пары праймеров были следующими: CD4, 5′-CTGATGTGGAAGGCAGAGAAGGATTC/5′-CAG CACGCAAGCCAGGAACACTGTCT; Egr-1, 5′-AATCCTCAAGGGGAGCCGAGCGAACA/5′ — GAGTAGATGGGACTGCTGCTGTCGTTGGA; N-Ras, 5′-GGTGGTGGTTGGAGCAGGTGGTGTTG/5′-CCATGGGGACATCATCAGAAT C TTTC; Egr-2, 5′-CCCCTTTGACCAGATGAACGGAGTGG/ 5′-TGGATGGCGGCGATAAGAATGCTGAA; Egr-3, 5′-CGACTCGGTAGCCCATTACAATCAGA/5′-GAGATGCCCGCAGTTGGAATAAGGAG; CD69, 5′-CTACCTGCAAGAATGAGTGGATTTCA/5′-TTTTGTGGTTCACGGACACGCACCTC.
Анализ изменения электрофоретической подвижности.

Олигонуклеотиды, содержащие консенсусный сайт Egr-1 (подчеркнутый), 5′-CCCGGCGCGGGGGCGATTTCGAGTCA и 5′-TGACTCGAAATCGCCC или перекрывающиеся сайты Egr-1/SP1 (сайт SP1 выделен жирным шрифтом) 5′-GGAGGAGCGGCG GGGGCG GGC 90G and CG ‘-CCGGCGCCCGCCCCCGC отжигали и метили в реакции заполнения с использованием [α 32 P]dCTP (ICN, Costa Mesa, CA) и фрагмента ДНК-полимеразы Кленова (GIBCO BRL), как описано ранее (26). Меченые олигонуклеотидные зонды отделяли от свободных нуклеотидов с помощью спин-колонок (Qiagen) и ∼3 × 10 9В каждой реакции связывания использовали зонд 0940 5 имп/мин (2–10 × 10 5 имп/мин/нг). Ядерные экстракты из 5 ? NP-40, 10 мкг/мл пепстатина, антипаина, химостатина, апротинина и 0,1 мкг/мл лейпептина, 0,5 мМ PMSF). После 5-минутной инкубации при 4°С ядра осаждали при 14000 об/мин в течение 10 с, однократно промывали 100 мкл буфера А, затем ресуспендировали в 50 мкл буфера В (20 мМ Hepes, рН 7,9)., 20% глицерина, 100 мМ KCl, 100 мМ NaCl, 0,2 мМ ЭДТА, 0,5 мМ PMSF, 0,5 мМ DTT, ингибиторы протеазы, как в буфере А). Ядерные экстракты замораживали в жидком азоте, оттаивали на льду, обрабатывали ультразвуком и хранили аликвотами при -70°С. Для анализа гель-сдвига 4–5 мкл ядерного экстракта инкубировали с радиоактивно меченым фрагментом ДНК в течение 20 мин при комнатной температуре в 20 мкл реакционной смеси, содержащей 1× связывающий буфер (20 мМ Hepes, pH 7,9, 50 мМ KCl, 5,0% глицерин, 1 мМ DTT, 1 мг/мл BSA, 0,1% Nonidet P-40 и 50 мкг/мл поли-dI-dC). Комплексы белок-ДНК отделяли от негибридизированного зонда в 6% неденатурирующих акриламидных гелях (Novex, Сан-Диего, Калифорния) в буфере 0,5× Трис борат-ЭДТА.
Ядерные экстракты замораживали в жидком азоте, оттаивали на льду, обрабатывали ультразвуком и хранили аликвотами при -70°С. Для анализа гель-сдвига 4–5 мкл ядерного экстракта инкубировали с радиоактивно меченым фрагментом ДНК в течение 20 мин при комнатной температуре в 20 мкл реакционной смеси, содержащей 1× связывающий буфер (20 мМ Hepes, pH 7,9, 50 мМ KCl, 5,0% глицерин, 1 мМ DTT, 1 мг/мл BSA, 0,1% Nonidet P-40 и 50 мкг/мл поли-dI-dC). Комплексы белок-ДНК отделяли от негибридизированного зонда в 6% неденатурирующих акриламидных гелях (Novex, Сан-Диего, Калифорния) в буфере 0,5× Трис борат-ЭДТА.
Индукция фактора транскрипции Egr-1 в клеточной линии тимоцитов DPK.
Клеточная линия DPK была получена из CD4 + 8 + лимфомы тимуса трансгенной мыши AND TCR (38, 45). Эта клеточная линия обладает фенотипом незрелых тимоцитов, включая экспрессию RAG-1,2 и TdT (41), и может быть запущена для дифференцировки в клетки CD4 + 8 − при активации голубиным пептидом цитохрома с и Е. k , несущие антиген-презентирующие клетки, или альтернативно по Е k , несущие эпителиальные клетки тимуса in vivo или in vitro в отсутствие антигена (38, 46). Хотя mAb против CD3 сами по себе являются относительно слабым индуктором дифференцировки клеток DPK, они активны при высоких концентрациях или в сочетании со стимуляцией вспомогательными молекулами (41, 47). Чтобы идентифицировать гены, участвующие в ранней фазе опосредованной TCR двойной положительной дифференцировки тимоцитов, мы использовали анализ репрезентативных различий для сравнения мРНК, полученной из клеток DPK, которые культивировали в присутствии или в отсутствие mAb против CD3ε (см. Материалы и методы). Такой подход позволил выделить Egr-1 ранний ген, кодирующий фактор транскрипции цинковых пальцев, в качестве одного из таких генов-кандидатов. Последующий анализ RT-PCR подтвердил, что клетки DPK экспрессируют небольшое количество мРНК Egr-1 перед активацией, но экспрессируют высокие уровни уже через 1 час после стимуляции анти-CD3ε mAb (фиг.
k , несущие антиген-презентирующие клетки, или альтернативно по Е k , несущие эпителиальные клетки тимуса in vivo или in vitro в отсутствие антигена (38, 46). Хотя mAb против CD3 сами по себе являются относительно слабым индуктором дифференцировки клеток DPK, они активны при высоких концентрациях или в сочетании со стимуляцией вспомогательными молекулами (41, 47). Чтобы идентифицировать гены, участвующие в ранней фазе опосредованной TCR двойной положительной дифференцировки тимоцитов, мы использовали анализ репрезентативных различий для сравнения мРНК, полученной из клеток DPK, которые культивировали в присутствии или в отсутствие mAb против CD3ε (см. Материалы и методы). Такой подход позволил выделить Egr-1 ранний ген, кодирующий фактор транскрипции цинковых пальцев, в качестве одного из таких генов-кандидатов. Последующий анализ RT-PCR подтвердил, что клетки DPK экспрессируют небольшое количество мРНК Egr-1 перед активацией, но экспрессируют высокие уровни уже через 1 час после стимуляции анти-CD3ε mAb (фиг. 1, —). Уровни мРНК Egr-1 остаются высокими по крайней мере в течение 5 часов, но снижаются примерно в пять раз через 1 день после первоначальной стимуляции (рис. 1, –). Чтобы установить, приводит ли индукция мРНК Egr-1 к ДНК-связывающей активности, мы провели анализ сдвига электрофоретической подвижности с использованием ядерных экстрактов, полученных из клеток DPK, активированных mAb против CD3ε. Ядерные экстракты из клеток DPK в отсутствие активации не имели детектируемых комплексов, связывающихся с олигонуклеотидным зондом, содержащим консенсусную связывающую последовательность Egr-1 (рис. 1, 9).0363 B ), что соответствует низкому уровню мРНК Egr-1. Напротив, в активированных клетках DPK наблюдался одиночный связывающий комплекс (рис. 1, B ). Этот комплекс имеет идентичную подвижность той, что получена при использовании рекомбинантного Egr-1, и суперсдвиги с антителом против Egr-1 (не показано). Индуцируемость ДНК-связывающей активности Egr-1 отличается от ДНК-связывающей активности Sp1, которая конститутивно экспрессируется в клетках DPK и не зависит от стимуляции (рис.
1, —). Уровни мРНК Egr-1 остаются высокими по крайней мере в течение 5 часов, но снижаются примерно в пять раз через 1 день после первоначальной стимуляции (рис. 1, –). Чтобы установить, приводит ли индукция мРНК Egr-1 к ДНК-связывающей активности, мы провели анализ сдвига электрофоретической подвижности с использованием ядерных экстрактов, полученных из клеток DPK, активированных mAb против CD3ε. Ядерные экстракты из клеток DPK в отсутствие активации не имели детектируемых комплексов, связывающихся с олигонуклеотидным зондом, содержащим консенсусную связывающую последовательность Egr-1 (рис. 1, 9).0363 B ), что соответствует низкому уровню мРНК Egr-1. Напротив, в активированных клетках DPK наблюдался одиночный связывающий комплекс (рис. 1, B ). Этот комплекс имеет идентичную подвижность той, что получена при использовании рекомбинантного Egr-1, и суперсдвиги с антителом против Egr-1 (не показано). Индуцируемость ДНК-связывающей активности Egr-1 отличается от ДНК-связывающей активности Sp1, которая конститутивно экспрессируется в клетках DPK и не зависит от стимуляции (рис.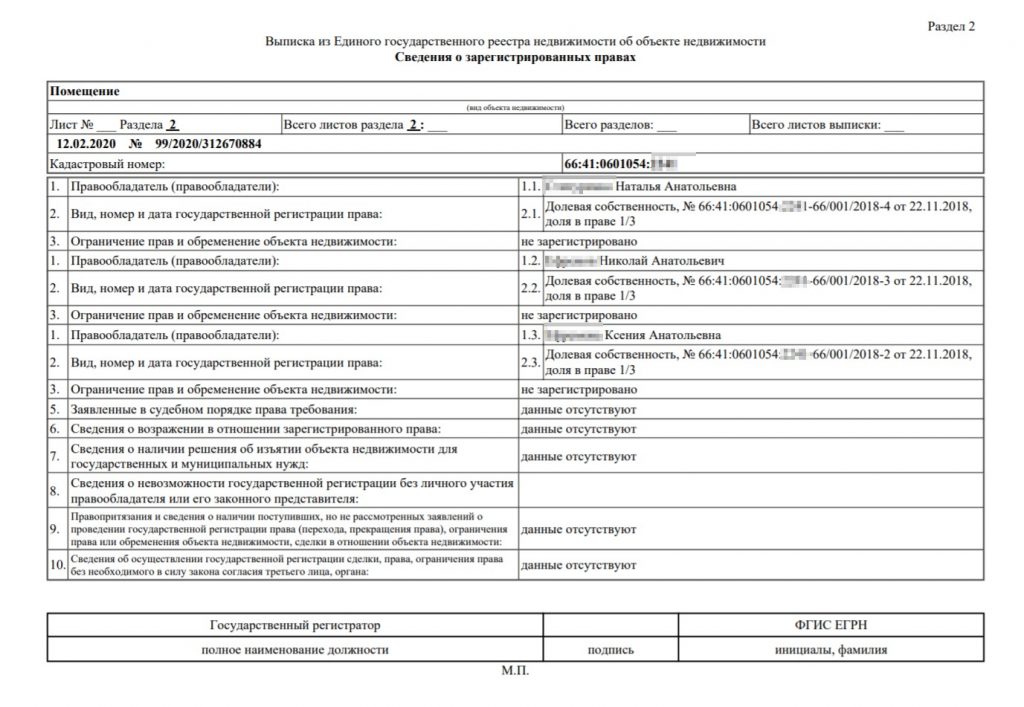 1 B ).
1 B ).
Быстрая и высокая экспрессия мРНК Egr-1 также наблюдается при активации клеток DPK пептидом цитохрома с голубя и трансфицированными антигенпрезентирующими клетками фибробластов (рис. 1, C ), мощным индуктором дифференцировки клеток DPK (38 ). В этом случае для количественного определения и сравнения уровней мРНК Egr-1 и CD4 использовали конкурентный анализ ПЦР. Экспрессию гена CD4 использовали для сравнения, поскольку экспрессия этого гена сохраняется на протяжении всей дифференцировки клеток DPK (38). Клетки DPK, активированные антигеном и антигенпрезентирующими клетками или анти-CD3ε mAb, экспрессируют в 20–30 раз более высокие уровни Egr-1, чем мРНК CD4 (рис. 1 9).0363 С ).
Egr-1 относится к семейству факторов транскрипции, которые имеют высококонсервативный домен связывания ДНК, состоящий из трех мотивов «цинковые пальцы». Чтобы определить, повышалась ли экспрессия генов других членов семейства при активации незрелых Т-клеток, мы исследовали экспрессию генов Krox-20, мышиного гомолога Egr-2 человека (16, 18) и Egr-3 (20) в клетки ДПК. Ни мРНК Krox20 (Egr-2), ни Egr-3 не были обнаружены с помощью ОТ-ПЦР в неактивированных клетках DPK, в то время как оба гена индуцировались после стимуляции mAb против CD3ε (рис. 2 9).0363 С ). Продукты ОТ-ПЦР Krox-20 (Egr-2) и Egr-3 секвенировали для подтверждения идентичности экспрессируемых генов.
Ни мРНК Krox20 (Egr-2), ни Egr-3 не были обнаружены с помощью ОТ-ПЦР в неактивированных клетках DPK, в то время как оба гена индуцировались после стимуляции mAb против CD3ε (рис. 2 9).0363 С ). Продукты ОТ-ПЦР Krox-20 (Egr-2) и Egr-3 секвенировали для подтверждения идентичности экспрессируемых генов.
Циклоспорин А ингибирует дифференцировку DPK и экспрессию гена Egr-2,3, но не Egr-1.
Наши результаты показали, что вовлечение TCR в незрелую двойную положительную клеточную линию DPK приводит к усилению регуляции членов семейства генов Egr . Однако оставалось определить, была ли индукция этих генов ниже сигнальных путей, которые необходимы для дифференцировки незрелых Т-клеток. Сообщалось, что циклоспорин А (CsA), мощный ингибитор кальцинейрина, блокирует положительный отбор (36, 48, 49).). Чтобы выяснить, зависит ли экспрессия гена Egr в двойных положительных клетках от активации кальциневрина, мы проверили способность CsA блокировать индукцию Egr в клетках DPK. Как и в случае с продукцией одиночных положительных тимоцитов, CsA ингибирует продуцирование CD4 одиночных положительных клеток DPK при активации антигеном и антигенпрезентирующими клетками (фиг. 2 B ). Понижающая регуляция CD8 клеточной поверхности уже через 24 ч после активации также блокируется CsA (не показано), что указывает на то, что кальциневрин участвует в инициации этого процесса, а не просто выживание единичных положительных клеток.
Как и в случае с продукцией одиночных положительных тимоцитов, CsA ингибирует продуцирование CD4 одиночных положительных клеток DPK при активации антигеном и антигенпрезентирующими клетками (фиг. 2 B ). Понижающая регуляция CD8 клеточной поверхности уже через 24 ч после активации также блокируется CsA (не показано), что указывает на то, что кальциневрин участвует в инициации этого процесса, а не просто выживание единичных положительных клеток.
Одним из самых ранних маркеров клеточной поверхности положительного отбора в тимусе (50–52) и в клетках DPK (38) является CD69. В активированных клетках DPK не наблюдалось значительного влияния CsA на раннюю индукцию мРНК CD69 (фиг. 2, C ). Через 24 часа после активации CsA частично снижал, но не устранял экспрессию CD69 на клеточной поверхности клеток DPK (фиг. 2, —). Однако к 3 дню поверхностная экспрессия CD69 в клетках DPK отсутствовала, что позволяет предположить, что кальциневрин может участвовать в поддержании, но не в индукции CD69. выражение. Таким образом, хотя ранняя индукция гена CD69 относительно устойчива к действию CsA, инициация дифференцировки в одну положительную клетку явно чувствительна к CsA. CsA, однако, мало влиял на индукцию мРНК Egr-1. Используя экспрессию CD4 для нормализации, клетки DPK, которые были активированы в присутствии CsA, экспрессировали примерно 70% уровней мРНК Egr-1, экспрессируемых клетками, активированными в отсутствие CsA (фиг. 2, C ). Напротив, CsA почти полностью блокирует индукцию мРНК Egr-2 (Krox20) и Egr-3 (рис. 2 9).0363 С ). Т.о., активация некоторых, но не всех членов семейства генов Egr происходит ниже сигнального пути, необходимого для положительного отбора.
выражение. Таким образом, хотя ранняя индукция гена CD69 относительно устойчива к действию CsA, инициация дифференцировки в одну положительную клетку явно чувствительна к CsA. CsA, однако, мало влиял на индукцию мРНК Egr-1. Используя экспрессию CD4 для нормализации, клетки DPK, которые были активированы в присутствии CsA, экспрессировали примерно 70% уровней мРНК Egr-1, экспрессируемых клетками, активированными в отсутствие CsA (фиг. 2, C ). Напротив, CsA почти полностью блокирует индукцию мРНК Egr-2 (Krox20) и Egr-3 (рис. 2 9).0363 С ). Т.о., активация некоторых, но не всех членов семейства генов Egr происходит ниже сигнального пути, необходимого для положительного отбора.
Дифференцировка DPK и экспрессия гена Egr заблокированы доминантным негативным мутантом p21Ras.
Было показано, что у трансгенных мышей, экспрессирующих доминантно-негативные мутанты p21ras или MEK, происходит ингибирование продукции единичных положительных тимоцитов (53, 54). Таким образом, мы стремились определить эффект блока передачи сигналов ras при экспрессии семейства Egr в двойных положительных клетках. Кроме того, поскольку положительный отбор может быть многоступенчатым процессом, из трансгенных экспериментов не было ясно, требуется ли активация ras для инициации перехода от двойного положительного к одиночному положительному или, альтернативно, для выживания тимоцитов или других более поздних событий.
Таким образом, мы стремились определить эффект блока передачи сигналов ras при экспрессии семейства Egr в двойных положительных клетках. Кроме того, поскольку положительный отбор может быть многоступенчатым процессом, из трансгенных экспериментов не было ясно, требуется ли активация ras для инициации перехода от двойного положительного к одиночному положительному или, альтернативно, для выживания тимоцитов или других более поздних событий.
Как показано на рис. 3, A , мРНК N-ras может быть обнаружена в нормальных тимоцитах, тимоцитах, полученных от мышей с дефицитом MHC (> 90% двойных положительных результатов, рис. 2, A ), и клетках DPK . Это согласуется с предыдущими сообщениями об экспрессии как N-ras, так и K-ras в дважды положительных тимоцитах (53). Чтобы изучить роль p21ras в экспрессии гена Egr в незрелых Т-клетках, ретровирусный перенос гена использовали для экспрессии доминантно-негативного мутанта p21 ras 9.0941, Haras N17 (с аспарагином, замещенным серином в положении 17) в клетках DPK. Этот мутант блокирует эндогенную функцию ras, конкурируя за белки обмена гуаниновых нуклеотидов, тем самым предотвращая образование комплексов ras-GTP (42). На рис. 3 B показана экспрессия Ha-ras N17 в четырех независимых клеточных трансформантах DPK, которые экспрессируют высокие уровни этого доминантно-негативного мутанта. Здесь описаны результаты, полученные с линией 17N4, хотя все четыре клеточные линии имели сходные ответы.
Этот мутант блокирует эндогенную функцию ras, конкурируя за белки обмена гуаниновых нуклеотидов, тем самым предотвращая образование комплексов ras-GTP (42). На рис. 3 B показана экспрессия Ha-ras N17 в четырех независимых клеточных трансформантах DPK, которые экспрессируют высокие уровни этого доминантно-негативного мутанта. Здесь описаны результаты, полученные с линией 17N4, хотя все четыре клеточные линии имели сходные ответы.
Одно из основных различий между клетками DPK и большинством популяций двойных положительных тимоцитов заключается в том, что клетки DPK быстро делятся. Учитывая потенциальную роль ras как протоонкогена, возможно, что блокирование p21ras-опосредованной передачи сигналов будет изменять рост клеток DPK и вторичную дифференцировку. Однако сверхэкспрессия rasN17 не влияла на рост клеток DPK и не наблюдалось каких-либо изменений в морфологии (не показано). Кроме того, как клетки DPK, так и клетки 17N4 проявляли сходную способность мобилизовать кальций после вовлечения TCR (не показано). В клетках Jurkat активация p21ras необходима и достаточна для индукции маркера ранней активации CD69.(55). Сходным образом, экспрессия rasN17 в клетках DPK устраняет поверхностную экспрессию CD69 при активации (Fig. 3, C ). Кроме того, анализ ОТ-ПЦР показал, что мРНК CD69 может быть обнаружена уже через 6 ч после активации антигеном в клетках дикого типа, но не в клетках 17N4 (не показано). Это контрастирует с эффектами CsA, которые мало влияли на раннюю индукцию CD69. В ответ на антиген клетки DPK переходят от двойного положительного к CD4 + 8 — одиночном положительному фенотипу. Подобно нормальным тимоцитам, потеря CD8 клеточной поверхности в клетках DPK представляет собой постепенный процесс, и переходные клетки с низким уровнем CD8 наблюдаются в ранние моменты времени (рис. 3, 9).0363 Д ). Продукция одиночных CD4-положительных клеток в ответ на антиген почти полностью ингибируется в клетках 17N4 даже в эти ранние моменты времени (рис. 3 D ).
В клетках Jurkat активация p21ras необходима и достаточна для индукции маркера ранней активации CD69.(55). Сходным образом, экспрессия rasN17 в клетках DPK устраняет поверхностную экспрессию CD69 при активации (Fig. 3, C ). Кроме того, анализ ОТ-ПЦР показал, что мРНК CD69 может быть обнаружена уже через 6 ч после активации антигеном в клетках дикого типа, но не в клетках 17N4 (не показано). Это контрастирует с эффектами CsA, которые мало влияли на раннюю индукцию CD69. В ответ на антиген клетки DPK переходят от двойного положительного к CD4 + 8 — одиночном положительному фенотипу. Подобно нормальным тимоцитам, потеря CD8 клеточной поверхности в клетках DPK представляет собой постепенный процесс, и переходные клетки с низким уровнем CD8 наблюдаются в ранние моменты времени (рис. 3, 9).0363 Д ). Продукция одиночных CD4-положительных клеток в ответ на антиген почти полностью ингибируется в клетках 17N4 даже в эти ранние моменты времени (рис. 3 D ). В совокупности эти результаты указывают на то, что передача сигналов p21ras необходима для инициации дифференцировки клеток DPK в одну CD4-позитивную клетку.
В совокупности эти результаты указывают на то, что передача сигналов p21ras необходима для инициации дифференцировки клеток DPK в одну CD4-позитивную клетку.
Определив, что дифференцировка клеток DPK зависит от активации ras, мы попытались определить, находится ли экспрессия семейства генов Egr ниже по течению от этого сигнального пути. Как показано на рис. 4, A , индукция мРНК Egr-1 в клетках DPK блокировалась экспрессией доминантно-негативного мутанта p21ras. Точно так же не было заметной индукции мРНК Egr-2 (Krox-20) или Egr-3 в этих клетках после стимуляции анти-CD3ε mAb (фиг. 4 B ). Это отличается от CsA, который по-разному влияет на экспрессию семейства Egr.
Egr-1 экспрессируется дважды положительными тимоцитами in vivo в результате распознавания MHC.
Сообщалось, что тимус новорожденных и взрослых крыс экспрессирует мРНК Egr-1 (37). Наши результаты в системе DPK показали, что экспрессия Egr-1 в нормальном тимусе может быть результатом положительного отбора. Чтобы решить эту проблему, мы использовали конкурентную ОТ-ПЦР для анализа Экспрессия гена Egr-1 в нормальных тимоцитах и в тимоцитах, полученных от мышей с двойным нокаутом гена, которые не экспрессируют молекулы MHC класса I или класса II (39, 40). Эти последние животные имеют нормальное количество двойных положительных тимоцитов, но не имеют одиночных положительных тимоцитов из-за неспособности незрелых тимоцитов, несущих TCR, подвергаться положительному отбору (фиг. 5, —). Свежевыделенные тимоциты животных с дефицитом MHC экспрессировали примерно в 10–20 раз меньше мРНК Egr-1, чем нормальные тимоциты (рис. 5, 9).0363 В ). Напротив, экспрессия гена CD4 была одинаковой в обеих группах животных, что согласуется с экспрессией этого корецептора на >90% тимоцитов как у нормальных, так и у мышей с дефицитом MHC (фиг. 5, —). Для анализа функциональной экспрессии белка Egr-1 были проведены анализы изменения подвижности геля с использованием ядерных экстрактов, полученных из свежевыделенных тимоцитов.
Чтобы решить эту проблему, мы использовали конкурентную ОТ-ПЦР для анализа Экспрессия гена Egr-1 в нормальных тимоцитах и в тимоцитах, полученных от мышей с двойным нокаутом гена, которые не экспрессируют молекулы MHC класса I или класса II (39, 40). Эти последние животные имеют нормальное количество двойных положительных тимоцитов, но не имеют одиночных положительных тимоцитов из-за неспособности незрелых тимоцитов, несущих TCR, подвергаться положительному отбору (фиг. 5, —). Свежевыделенные тимоциты животных с дефицитом MHC экспрессировали примерно в 10–20 раз меньше мРНК Egr-1, чем нормальные тимоциты (рис. 5, 9).0363 В ). Напротив, экспрессия гена CD4 была одинаковой в обеих группах животных, что согласуется с экспрессией этого корецептора на >90% тимоцитов как у нормальных, так и у мышей с дефицитом MHC (фиг. 5, —). Для анализа функциональной экспрессии белка Egr-1 были проведены анализы изменения подвижности геля с использованием ядерных экстрактов, полученных из свежевыделенных тимоцитов. В соответствии с паттерном экспрессии мРНК Egr-1 активность связывания ДНК Egr-1 была обнаружена в ядерных лизатах тимоцитов, полученных от животных дикого типа, но не дефицитных по MHC (рис. 5, 9).0363 С ). Этот комплекс имеет идентичную подвижность, полученную с рекомбинантным Egr-1, и суперсдвиги с антителом против Egr-1 (рис. 5 C ).
В соответствии с паттерном экспрессии мРНК Egr-1 активность связывания ДНК Egr-1 была обнаружена в ядерных лизатах тимоцитов, полученных от животных дикого типа, но не дефицитных по MHC (рис. 5, 9).0363 С ). Этот комплекс имеет идентичную подвижность, полученную с рекомбинантным Egr-1, и суперсдвиги с антителом против Egr-1 (рис. 5 C ).
Эти результаты свидетельствуют о том, что активация гена Egr-1 произошла во время MHC-зависимой селекции двойных положительных тимоцитов. Однако данные также согласовывались с возможностью того, что Egr-1 активировался только в зрелых единичных положительных тимоцитах после положительного отбора. Чтобы различить эти возможности, мы проанализировали экспрессию Egr-1 в очищенных двойных положительных тимоцитах, полученных от нормальных мышей. Учитывая быструю индукцию мРНК Egr-1 в клетках DPK (Fig. 1), можно было бы предсказать, что если Egr-1 индуцируется во время тимусной селекции, он будет экспрессироваться до потери двойного положительного фенотипа. Это оказалось так, и значительное количество мРНК Egr-1 было обнаружено в дважды положительных тимоцитах, выделенных методом клеточного сортинга (рис. 6, 9).0363 А ). Изолированные двойные положительные тимоциты также проявляли ДНК-связывающую активность Egr-1, как и CD4 одиночные положительные тимоциты (не показано). Чтобы продемонстрировать, что ген Egr-1 может быть индуцирован активацией TCR в дважды положительных тимоцитах, тимоциты, полученные от мышей с дефицитом MHC, стимулировали иммобилизованным анти-CD3ε mAb. Как показано на фиг. 6 B , активация TCR приводит к быстрой активизации мРНК Egr-1 в этих клетках. Напротив, Egr-1 не активируется в тимоцитах, полученных от мышей с дефицитом MHC, при обработке в культуре дозами дексаметазона или ионизирующего излучения, которые вызывают гибель клеток (не показано).
Это оказалось так, и значительное количество мРНК Egr-1 было обнаружено в дважды положительных тимоцитах, выделенных методом клеточного сортинга (рис. 6, 9).0363 А ). Изолированные двойные положительные тимоциты также проявляли ДНК-связывающую активность Egr-1, как и CD4 одиночные положительные тимоциты (не показано). Чтобы продемонстрировать, что ген Egr-1 может быть индуцирован активацией TCR в дважды положительных тимоцитах, тимоциты, полученные от мышей с дефицитом MHC, стимулировали иммобилизованным анти-CD3ε mAb. Как показано на фиг. 6 B , активация TCR приводит к быстрой активизации мРНК Egr-1 в этих клетках. Напротив, Egr-1 не активируется в тимоцитах, полученных от мышей с дефицитом MHC, при обработке в культуре дозами дексаметазона или ионизирующего излучения, которые вызывают гибель клеток (не показано).
Подобно результатам, полученным с клетками DPK, мРНК Krox-20 (Egr-2) и Egr-3 также экспрессируются в нормальных тимоцитах, в то время как экспрессия этих генов не обнаруживается в тимоцитах, полученных от животных с дефицитом MHC (рис. 7). Эти результаты согласуются с координированной активацией семейства факторов транскрипции Egr во время тимусной селекции.
Для определения локализации в тимусе клеток, экспрессирующих белок Egr-1, тонкие срезы нормального тимуса окрашивали специфической антисывороткой против пептида Egr-1 (рис. 8). Было обнаружено, что изолированные клетки, разбросанные по всей коре, экспрессируют Egr-1. Не наблюдалось обогащения клеток, экспрессирующих Egr-1, в субкапсулярной области или кортико-медуллярном соединении. Эти результаты согласуются с индукцией Egr-1 в кортикальных двойных положительных тимоцитах в результате вовлечения TCR. Большинство, но не все, медуллярные клетки также окрашивались, что указывает на то, что белок Egr-1 также экспрессируется в значительном количестве единичных положительных тимоцитов (см. ниже). Напротив, в коре тимуса мышей с дефицитом MHC окрашивалось мало клеток, и в целом окрашивание было слабым даже в медуллярных областях (рис. 8).
Экспрессию Egr-1 в субпопуляциях тимоцитов дополнительно анализировали с помощью FACS ® с использованием трехцветного поверхностного окрашивания в сочетании с четвертым цветом для внутреннего окрашивания (рис. 9). Используя этот метод, можно было обнаружить подмножество тимоцитов, окрашенных антисывороткой против Egr-1. Эта субпопуляция включала большинство одиночных положительных тимоцитов и ~10% двойных положительных тимоцитов. Эти результаты согласуются с тем фактом, что изолированные кортикальные тимоциты и большинство медуллярных тимоцитов окрашиваются этой антисывороткой против Egr-1. Интересно, что почти все CD69 + или CD3 hi тимоциты также окрашивались на Egr-1, в то время как ~10% тимоцитов окрашивались на Egr-1, но были CD3 lo или CD69 — (рис. 9). Среди дважды положительных тимоцитов >90% клеток, окрашенных на Egr-1, также были CD3 -10941 или CD69 —. Большинство двойных негативных тимоцитов также окрашиваются на Egr-1, хотя специфичность этого окрашивания еще предстоит подтвердить другими способами.
Используя анализ репрезентативных различий и систему культивирования DPK, мы стремились идентифицировать гены, которые играют роль на самых ранних стадиях дифференцировки двойных положительных тимоцитов. Egr-1 , ранний ген, кодирующий фактор транскрипции цинковых пальцев, был идентифицирован как один из таких кандидатов. Мы продемонстрировали, что ген Egr-1 быстро индуцируется после опосредованной TCR стимуляции двойной положительной клеточной линии DPK или дважды положительных тимоцитов. У нормальных мышей активность связывания мРНК, белка и ДНК Egr-1 экспрессируется свежевыделенными тимоцитами, включая двойную положительную субпопуляцию. Напротив, тимоциты, полученные от мышей с дефицитом MHC, которые демонстрируют нормальный паттерн развития через двойную положительную стадию, но которые не подвергаются положительной или отрицательной селекции, экспрессируют примерно в 10-20 раз более низкие уровни мРНК Egr-1, чем дикие. типа мыши. Это коррелирует с потерей обнаруживаемой ДНК-связывающей активности Egr-1 и низким уровнем окрашивания антител против Egr-1 в тимоцитах, полученных от мышей с дефицитом MHC. Кроме того, экспрессия Egr-1 также низка в тимоцитах, полученных от мышей с нокаутом α-цепи TCR (не показано), демонстрируя, что для высокого уровня Egr-1 требуется взаимодействие TCR-MHC, а не, например, взаимодействие корецептор-MHC. экспрессия в тимусе. Эти результаты противоречат экспрессии активности связывания ДНК Sp1, которая конститутивно экспрессируется в двойных положительных тимоцитах (56) и клетках DPK до и после активации (рис. 1).
Возникает вопрос, подвергаются ли тимоциты, экспрессирующие Egr-1, положительной селекции. Положительный отбор двойных положительных тимоцитов инициируется взаимодействием с эпителиальными клетками, несущими MHC, которые находятся в коре тимуса (57). Если индукция Egr-1 в двойных положительных клетках является следствием положительного отбора, то можно было бы ожидать, что она также имеет место в коре. Было обнаружено, что это так, и окрашивание срезов тимуса на Egr-1 выявило разбросанные по коре тимоциты, экспрессирующие белок. Egr-1 также экспрессируется многими медуллярными (рис. 8) и единичными положительными тимоцитами (рис. 9).). Неизвестно, поддерживается или восстанавливается экспрессия Egr-1 на этих более зрелых тимоцитах, но активация Egr-1 связана как с ранней активацией, так и с поздней дифференцировкой в других системах (28).
В то время как мы не можем напрямую определить судьбу экспрессирующих Egr-1 клеток в коре, тимоциты с повышенной экспрессией CD3 или CD69, включая CD4 + 8 lo переходные тимоциты, также имеют повышенную регуляцию Egr-1 (рис. 9). ). Этот паттерн экспрессии в сочетании с нашими результатами в системе DPK убедительно свидетельствует о том, что Egr-1 индуцируется во время положительного отбора. Приблизительно 10% от общего числа тимоцитов, большинство из которых также являются дважды положительными тимоцитами, также окрашенными на Egr-1 с помощью FACS ® , но были CD3 lo или CD69 — (рис. 9). Это может отражать быструю индукцию Egr-1 во время селекции перед активацией TCR или CD69. Наблюдаемая быстрая активация Egr-1 в клетках DPK и тимоцитах в результате вовлечения TCR определенно согласуется с этой возможностью. Фактически, в линии CD4 активация TCR происходит после стадии двойного позитива и совпадает с подавлением корецептора CD8 (58). Таким образом, экспрессия Egr-1 может представлять собой один из самых ранних маркеров положительного отбора в тимусе. Альтернативно, некоторые тимоциты, которые экспрессируют Egr-1 на стадии двойного позитива, могут не продвигаться в своей программе дифференцировки или находиться в процессе негативного отбора (см. ниже).
Хотя экспрессия Egr-1 связана с дифференцировкой тимоцитов, может ли активация этого фактора транскрипции также происходить во время негативного отбора? В подтверждение этого мы показали, что двойные положительные тимоциты, стимулированные анти-CD3-антителом, активируют мРНК Egr-1. Сигналы, обеспечиваемые перекрестным связыванием CD3, не вызывают положительной селекции изолированных нормальных тимоцитов, но могут вызывать гибель клеток в сочетании с дополнительными сигналами (59). Следовательно, экспрессия Egr-1 может не ограничиваться положительным отбором, но также может индуцироваться во время отрицательного отбора. Если действительно высокий уровень экспрессии Egr-1 является результатом как положительного, так и отрицательного отбора, то паттерн экспрессии этого транскрипционного фактора в коре может выявить степень специфического участия TCR в нормальном тимусе.
Хотя Egr-1 может индуцироваться во время отрицательного отбора, данные свидетельствуют о том, что индукция этого транскрипционного фактора не связана с гибелью клеток тимоцитов как таковая. Хотя апоптотические тимоциты могут быть обнаружены в коре головного мозга нормальных мышей, у мышей с двойным нокаутом MHC не наблюдается снижения частоты таких клеток (60). Предположительно, гибель клеток в последнем случае является результатом гибели от пренебрежения, а не отрицательного отбора. Напротив, наблюдается резкое снижение активности связывания мРНК, белка и ДНК Egr у мышей с нокаутом MHC, что позволяет предположить, что гибель тимоцитов в результате пренебрежения не связана с активацией Egr-1. Нам также не удалось обнаружить индукцию мРНК Egr-1 в тимоцитах с нокаутом MHC, подвергнутых ионизирующему излучению или кортикостероидам в дозах, вызывающих гибель клеток. Это дополнительно демонстрирует, что высокий уровень экспрессии мРНК Egr-1 в тимусе специфически связан с активацией TCR.
Маловероятно, что события селекции, опосредованные TCR, являются единственным источником экспрессии гена Egr-1 в тимусе, поскольку некоторое количество мРНК Egr-1 может быть обнаружено у животных с дефицитом MHC. Двойные положительные тимоциты могут конститутивно экспрессировать ген на очень низких уровнях, потому что мРНК Egr-1 также может быть обнаружена в клетках DPK с помощью RT-PCR, если используется достаточное количество кДНК. Поскольку на ДНК-связывающую активность Egr-1 может влиять фосфорилирование (61), как транскрипционные, так и посттрансляционные механизмы могут способствовать усилению функционального Egr-1 в незрелых тимоцитах. Кроме того, мы наблюдали окрашивание двойных негативных тимоцитов с использованием антисыворотки против Egr-1 и FACS 9.Анализ 0940 ® (рис. 9). Поскольку двойные негативные клетки, как правило, имеют более высокий фон окрашивания для многих антител, этот результат необходимо подтвердить. Однако одна интригующая возможность заключается в том, что Egr-1 также индуцируется активацией pre-TCR (62) и играет роль в дифференцировочных и/или пролиферативных событиях, которые участвуют в переходе тимоцитов от двойного отрицательного к двойному положительному.
Egr-1 принадлежит к семейству регуляторов транскрипции, которые содержат близкородственные ДНК-связывающие домены, распознающие похожие мотивы последовательностей. Однако, несмотря на очень сходные ДНК-связывающие домены, были обнаружены различия в способности членов семейства Egr взаимодействовать с данной промоторной областью (63). Таким образом, вопрос о степени функционального совпадения между членами семьи остается открытым. Сообщалось, что Egr-1, 2, 3 и 4 активируются при активации Т-лимфоцитов человека (19)., 64). Мы обнаружили, что Egr-1, 2 и 3 также координированно активируются в тимусе, а экспрессия мРНК для всех трех членов семейства зависит от экспрессии MHC. Несколько неожиданно, учитывая эти результаты, в анализах гель-сдвига с использованием ядерных лизатов тимоцитов или DPK обнаруживается только один связывающий комплекс. Этот комплекс работает идентично комплексу, полученному с рекомбинантным Egr-1, и комплекс суперсдвигается с антителом против Egr-1. Возможно, что невозможность обнаружения комплексов, содержащих Egr-2 и Egr-3, связана с более низкими уровнями экспрессии этих транскрипционных факторов (19). ) или к конкретным условиям связывания анализа.
Положительный и отрицательный отбор представляют собой сложные процессы, включающие активацию клеток, их дифференцировку и выживание клеток. Как конкретно экспрессия Egr влияет на эти события, неизвестно. У мышей с нокаутом Egr-1 нет грубых аномалий лимфоцитов, включая продукцию зрелых Т-клеток (65), в то время как мыши с нокаутом Egr-2 умирают в младенчестве из-за аномалий заднего мозга (66). Есть ли более тонкие изменения в развитии Т-клеток или функции Т-клеток у мышей с нокаутом Egr-1, или компенсаторные эффекты других членов семьи, еще предстоит определить. Критические гены-мишени для регуляции членами семейства Egr в контексте активации Т-клеток или развития тимоцитов также неизвестны, хотя Egr-1 участвует в регуляции IL-2 (63), фактора некроза опухоли (67), ICAM -1 (68) и CD44 (69) гены. Промотор гена CD69 может также иметь сайт связывания Egr-1 (70). Также сообщалось, что Egr-1 регулирует отсроченную раннюю экспрессию другого немедленно раннего гена, nur77 (71), который сам участвует в гибели клеток тимоцитов (72).
Несмотря на нехватку генов-мишеней, которые, как известно, регулируются Egr-1, существует общая тема, которая возникла в отношении его механизма действия. Sp1, который имеет гомологию с Egr-1 в домене цинковых пальцев, связывает GC-богатую, но отличную от Egr-1 последовательность. Сайты Egr-1 и Sp1 часто перекрываются в промоторах генов, что позволяет конкурировать. Недавно было показано, что индукция Egr-1 в результате повреждения эндотелиальных клеток может привести к замещению конститутивно экспрессированного Sp1, что приводит к активации генов, участвующих в заживлении ран или росте клеток (27). Аналогичное замещение Sp1 на Egr-1 может также играть роль в активации гена IL-2 (63), а также в регуляции Egr-1 сам ген (73). Как показано здесь, поскольку Sp1 конститутивно экспрессируется в двойных положительных тимоцитах, тогда как Egr-1 индуцируется, подобный процесс может происходить в тимусе. Приведет ли результат замещения Sp1 к Egr-1 в контексте развития Т-клеток к активации или репрессии генов, и как это повлияет на развитие Т-клеток, необходимо дождаться идентификации специфических генов-мишеней для Egr-1 в тимусе.
Если действительно имеет место индукция членов семейства Egr во время как отрицательного, так и положительного отбора, это не означает, что эти транскрипционные факторы играют одинаковые роли в этих различных событиях отбора. Чтобы решить эту проблему, мы задались вопросом, как сигнальные пути влияют как на двойную положительную клеточную дифференцировку, так и на экспрессию семейства Egr. Ранее было показано, что экспрессия доминантно-негативных ras или MEK у трансгенных животных ингибирует появление единичных позитивных тимоцитов (53, 54). Используя систему DPK, мы показали здесь, что доминантно-отрицательный ras также ингибирует инициацию перехода этих клеток от двойного положительного к одиночному положительному. Это коррелирует с ингибированием индукции Egr-1, 2 и 3. Также было показано, что индукция Egr-1 в В-клетках зависит от ras (74). Тот факт, что негативный отбор (53) и клеточная гибель клеток DPK (Shao, H. and J. Kaye, неопубликованные наблюдения) не нарушается в условиях, которые нарушают передачу сигналов ras и экспрессию Egr, предполагает, что семейство Egr вряд ли играет роль. обязательную роль в отрицательном отборе.
Мы также исследовали влияние ингибитора кальциневрина CsA на эту систему. Ранее было показано, что CsA ингибирует продукцию единичных положительных тимоцитов in vivo (36, 48) и in vitro (49). CsA также блокирует переход клеток DPK от двойного положительного к одиночному положительному. Однако, в отличие от эффектов доминантно-негативного ras, влияние CsA на раннюю индукцию CD69 было минимальным. Было обнаружено, что индукция Egr-1 устойчива к CsA, в то время как экспрессия Egr-2 и Egr-3 чувствительна к CsA. Также сообщалось, что индукция Egr-1 в зрелых Т-лимфоцитах человека является резистентной к CsA (75), в то время как индукция Egr-3 оказалась чувствительной к CsA (64). Сходным образом, индукция Egr-2, но не Egr-1 в мышиной линии В-клеток ингибируется CsA (35). Эта экспрессия Egr-2 и 3 требует сигнальных путей как ras, так и кальцинейрина, что напоминает аналогичное двойное требование для активации гена IL-2 (76, 77), и координированная активность этих сигнальных путей может быть общей чертой многих аспектов. активации как зрелых Т-клеток, так и дифференцировки незрелых тимоцитов. Имеются противоречивые сообщения о том, блокирует ли CsA отрицательный отбор (36, 48, 49)., 78), хотя тот факт, что CsA блокирует положительный отбор, безусловно, является усложняющим фактором. Тем не менее, ясно, что семейство транскрипционных факторов Egr специфически индуцируется активацией TCR в незрелых тимоцитах и лежит ниже двух известных путей, которые необходимы для положительного отбора.
Авторы благодарят N. Mackman и G. Parry за помощь в анализе сдвига геля, E. Adamson и G. Parry за рекомбинантный Egr-1, C. Der за конструкцию ras17N и J. Sprent за мышей с дефицитом MHC. Мы также благодарим докторов. Дж. Спрент и А. Фини за критический обзор этой рукописи. Это рукопись 10220-IMM из Научно-исследовательского института Скриппса.
Эти исследования были поддержаны грантами Национальных институтов здравоохранения AI33219 и AI31231 для J. Kaye.
1
Sukhatme
VP
,
Kartha
S
,
Toback
FG
,
Taub
R
,
Hoover
RG
,
Tsai-Morris
C-H
Новый ген реакции раннего роста, быстро индуцируемый митогенами фибробластов, эпителиальных клеток и лимфоцитов
Онкоген Рез
1987
1
343
355
[В паблике]
2
Sukhatme
VP
,
Cao
X
,
Chang
LC
,
Tsai-Morris
C-H
,
Stamenkovich
D
,
Феррейра
PCP
,
Коэн
DR
,
Edwards
SA
,
Shows
TB
,
Curran
T
et al
A zinc finger-encoding gene coregulated with c-fos during growth and differentiation, and после клеточной деполяризации
Клетка
1988
53
37
43
[В паблике]
3
Кристи
Б
,
Nathans
LF
,
Nathans
D
A gene activated mouse 3T3 cells by serum growth factors encodes a protein with “zinc fingers” sequences
Proc Natl Acad Sci USA
1988
85
7857
7861
[В паблике]
4
Lemaire
P
,
Revelant
O
,
Bravo
R
,
Charnay
P
Two mouse genes encoding potential transcription factors with identical DNA-binding domains are activated by growth factors in cultured cells
Proc Natl Acad Sci USA
1988
85
4691
4695
[В паблике]
5
Lim
RW
,
Varnum
BC
,
Herschman
2 HR0003
Клонирование последовательностей первичного ответа, индуцированных тетрадеканоил-фарболовым эфиром, и их экспрессия в швейцарских клетках 3T3, привлеченных к плотности и непролиферативному варианту
1987
1
263
9000 2703
10003
263
9000 2703
[В паблике]
6
Milbrandt
J
Ген, индуцированный фактором роста нервов, кодирует возможный фактор транскрипции
Science (Wash DC)
1987
238
797
799
[В паблике]
7
Irving
SG
,
June
CH
,
Zipfel
PF
,
Siebenlist
U
,
Kelly
K
Mitogen-induced гены подвержены множественным путям регуляции на начальных стадиях активации Т-клеток
Mol Cell Biol
1989
9
1034
1040
[В паблике]
8
Cao
X
,
Koski
RA
,
Gashler
A
,
McKiernan
M
,
Morris
CF
,
Гаффни
Р
,
Хейс
РВ
,
Сукхатме
VP
Идентификация и характеристика продукта гена EGR-1, ДНК-связывающий белок цинкового пальца цинкового пальца, индуцированный дифференцировкой и сигналами роста
моль-клеточный биол
1990
10 0003
1931
1939
1931
9000 1939
[В паблике]
9
Christy
B
,
Nathans
D
ДНК-связывающий сайт белка, индуцируемого фактором роста Zif268
USA Sci Natl
Proc0003
1989
86
8737
8741
[В паблике]
10
Pavletich
NP
,
PABO
CO
ЗИНК-ДНК: Кристаллическая структура комплекса ZIF268
292
Science (Wash DC)
19
1
2
2
2
2
2
292
.
809
817
[В паблике]
11
Свирнофф
AH
,
Milbrandt
J
ДНК-связывающая специфичность NGFI-A и связанных с ним факторов транскрипции цинк
Моль-клеточный биол
1995
15
2275
2287
15
2275
2287
15
2275
2287
[В паблике]
12
Gashler
AL
,
Swaminathan
S
,
Sukhatme
VP
Новая репрессионная модуля и обширная активация Domain, и A Bip’s Bip’s Defination Signization. фактор ранней транскрипции EGR-1: прототип семейства факторов транскрипции 9 с цинковыми пальцами0003
Cell Biol
1993
13
4556
4571
13
Russo
MW
,
Matheny
C
,
Milbrandt
J
Transcriptional activity of the zinc палец NGFI-A находится под влиянием его взаимодействия с клеточным фактором
Molec Cell Biol
1993
13
6858
6865
[В паблике]
14
Carman
JA
,
Monroe
JG
Белок EGR1 содержит дискретный транскрипционный домен
DANA DNA DANNA. 14
581
589
[В паблике]
15
Руссо
МВт
,
Севецон
BR
,
Milbrandt
J
Идентификация NAB1, репрессора NGFI-A-и Krox20-опосредованного транскрипции
Proc Natl SCI USA
1995
9000 2
6873
677777777777777777777777777777777777777777777777777777777777тели
70003777777777777777777777777777777777777777777777777777777777777777Н7
[В паблике]
16
Chavrier
P
,
Zerial
M
,
Lemaire
P
,
Almendral
0002 J
,
Bravo
R
,
Charnay
P
A Gene, кодирующий белок с помощью цинковых пальцев, активируется во время G0/G1 Transsition в культурных клетках
Embo (Eur Mal Orarge)
9000 2
9000 2
9000 2 9000 2
. 1988
7
29
35
[В паблике]
17
Шаврье
P
,
Janssen-Timmen
U
,
Mattei
M-G
,
нулевой
M
,
Bravo
R
,
Charnay
P
Структура, Chromosome Location, и Arxperive Gene Gene Gene-Fingere. продукты гена и корегуляция с протоонкогеном c-fos
Mol Cell Biol
1989
9
787
797
[В паблике]
18
Joseph
LJ
,
Le Beau
MM
,
Jamieson
GA
Jr,
Acharya
S
,
Shows
TB
,
Rowley
JD
,
Sukhatme
VP
Молекулярное клонирование, секвенирование и картирование EGR2, гена реакции раннего роста человека, кодирующего белок со структурой «цинк-связывающего пальца»
Proc Natl Acad Sci USA
1988
85
7164
7168
[В паблике]
19
Zipfel
PF
,
Irving
SG
,
Kelly
k
,
Siebenlist
,
Siebenlist
,
Siebenlist
,
. клетки
Mol Cell Biol
1989
9
1041
1048
[В паблике]
20
Patwardhan
S
,
Gashler
A
,
Siegel
MG
,
Chang
LC
,
Joseph
LJ
,
Шоу
ТБ
,
Ле Бо
ММ
,
Сукхатме
VP
EGR3, новый член семейства генов Egr, кодирующих факторы ранней транскрипции
[В паблике]
21
Crosby
SD
,
Puetz
JJ
,
Simburger
KS
,
Fahrner
TJ
,
Milbrandt
J
Ген раннего отклика NGFI-C кодирует активатор транскрипции цинкового пальца и является членом семейства связывающих белков GCGGGGGCG (GSG)
Mol Cell Biol
11 0003
3835
3841
[В паблике]
22
Muller
H-J
,
Skerka
C
,
Bialonski
A
,
Zipel
A
,
Zipel
,
,
A
,
,
0002 PF
Клон pAT 133 идентифицирует ген, который кодирует другой человеческий член класса генов, индуцированных фактором роста, с почти идентичными доменами цинковых пальцев
[В паблике]
23
Гашлер
A
,
Сукхатме
VP
Белок ответа на ранний рост 1 (Egr-1): прототип семейства факторов транскрипции «цинковые пальцы»
Prog Nuc Acid Res
1995
50
191
224
[В паблике]
24
NGUYEN
HQ
,
Hoffman-Liebermann
B
,
Liebermann
DA
Трансцепционный фактор Zinc Film
Сотовый
1993
72
197
209
[В паблике]
25
Krishnaraju
K
,
Nguyen
HQ
,
Liebermann
DA
,
Hoffman
DA
,
DA
,
DA
,
DA
,
DA
,
9000. гемопоэтические клетки
Molec Cell Biol
1995
15
5499
5507
[В паблике]
26
Cui
M-Z
,
Parry
GCN
,
Oeth
P
,
Larson
H
,
Smith
M
,
Huang
R-P
,
Adamson
ED
,
Mackman
N
Транскрипционная регуляция гена тканевого фактора опосредована клетками эпителия человека Spli-1 и EGR10003
J Biol Chem
1996
271
2731
2739
[В паблике]
27
KHACHIGIAN
LM
,
Lindner
V
,
Williams
AJ
,
Collins
T.
,
Collins
T.
,
. при повреждении сосудов
Science (Wash DC)
1996
271
1427
1431
[В паблике]
28
Sukhatme
VP
Ранние события транскрипции в росте клеток: семейство EGR
J Am Soc Nephrol
1990
1
859
866
[В паблике]
29
Сейферт
ВЛ
,
Сукхатме
ВП
,
Монро
3 Дж
0003
Дифференциальная экспрессия гена, кодирующего цинковые пальцы, в ответ на положительную и отрицательную передачу сигналов через рецепторный иммуноглобулин в мышиных В-лимфоцитах
[В паблике]
30
Seyfert
VL
,
McMahon
S
,
Glenn
W
,
CAO
W
,
CAO
W
,
CAO
X
,
Sukhatme
VP
,
Monroe
J
Egr-1 expression in surface Ig-mediated B cell activation — Kinetics and association with protein kinase C activation
J Immunol
1990
145
3647
3653
[В паблике]
31
Маковер
D
,
Кадди
М
,
Yum
S
,
Bradley
K
,
Alpers
J
,
Sukhatme
V
,
Reed
JC
Phorbol ester-mediated inhibition of growth и регуляция экспрессии протоонкогена в линии Т-клеточного лейкоза человека JURKAT
Онкоген
1991
6
455
460
[В паблике]
32
Perez-Castillo
AP
,
Pipaon
C
,
Garcia
I
,
Alemany
I
,
Alemany
IM. пролиферация
J Biol Chem
1993
268
19445
19450
[В паблике]
33
Сейферт
ВЛ
,
McMahon
SB
,
Glenn
WD
,
Yellen
AJ
,
Sukhatme
VP
,
Cao
XM
,
Monroe
JG
Метилирование немедленно-раннего индуцируемого гена как механизм индукции толерантности В-клеток0003
[В паблике]
34
KLAUS
SJ
,
Phillips
NE
,
Parker
DC
Эффекты IL-4 и FC GAMMA Antagement II Antagement на EGR-1. методом кросслинкинга мембранного иммуноглобулина
Molec Immunol
1993
30
1553
1558
[В паблике]
35
GOTSCHALK
AR
,
JOSEPH
LJ
,
Quintans
J
FCGRII. Иммунол
1994
152
2115
2122
[В паблике]
36
Дженкинс
МК
,
Шварц
РХ
,
Pardoll
DM
Влияние циклоспорина A на развитие Т-клеток и делеция клонального
[В паблике]
37
Watson
MA
,
Milbrandt
J
Экспрессия генов NGFI-A и NGFI-B, регулируемых фактором роста нервов 9 900 2 Развитие 90 развивающихся крыс0002 1990
110
173
183
[В паблике]
38
Kaye
J
,
Ellenberger
DE
Differentiation of an immature T cell line: a model of thymic positive selection
Cell
1992
71
423
435
[В паблике]
39
Грусби
МДж
,
Johnson
RS
,
Papaioannou
VE
,
Glimcher
LH
Depletion of CD4+T cells in major histocompatibility complex class II-deficient mice
Science (Wash DC)
1991
253
1417
1420
[В паблике]
40
Зийлстра
М
,
Бикс
М
,
Simister
NE
,
Loring
JM
,
Raulet
DH
,
Jaenisch
R
Beta-microglobulin deficient mice lack CD4-8+cytolytic T cells
Nature (Лондон)
1990
344
742
746
[В паблике]
41
ДеКонинг
J
,
Карбон
F
,
Kaye
J
Контраст между опосредованной дифференцировкой CD4+CD8+CD8+CD8+T-клеток: последствия для линии
Int Immunol
1995
7
9000 541 541
549
[В паблике]
42
Quilliam
LA
,
Kato
K
,
Rabun
км
,
0002 HISAKA
мм
,
HUFF
SY
,
Campbell-Burk
S
,
Der
CJ
Идентификация идентификации остатков для RAS-INIMITIRITITIRITIRITION и RAS-INIFITIRITER (17N) Der Der). для взаимодействия ras с факторами обмена гуаниновых нуклеотидов
Molec Cell Biol
1994
14
1113
1121
[В паблике]
43
Kuhlman
P
,
Moy
VT
,
Lollo
BA
,
Brian
AA
ОБЩЕСТВЕННАЯ ФУНЦИАЛЬНА Дж Иммунол
1991
146
1773
1782
[В паблике]
44
Хубанк
М
,
Шац
DG
Выявление различий в экспрессии мРНК с помощью репрезентативного анализа различий кДНК
Nuc Acids Res
1994
22
5640
5649
1994
[В паблике]
45
Kaye
J
,
Hsu
M-L
,
Sauron
M-E
,
Jameson
SC
,
Gascoigne
NRJ
,
Hedrick
SM
Selective development of CD4+T cells in transgenic mice expressing a class II MHC-restricted antigen receptor
Nature (Lond)
1989
341
746
749
[В паблике]
46
Пуарье
G
,
Lo
D
,
Рейли
3 9 9
CR0002 кей
j
Дискриминация между эпителиальными клетками тимуса и периферическим антигеном, представляющими клетки, при индукции незрелой дифференцировки Т -клеток
Иммунитет
1994
1
385
391
385
391
[В паблике]
47
ДеКонинг Дж. и Дж. Кэй. Требования к дифференцировке незрелой линии CD4+8+ Т-клеток. Дев. Иммунол. Под давлением.
48
Гао
ЕК
,
LO
D
,
Cheney
R
,
Kanagawa
,
Sprent
J
,
Sprent
J
,
. Природа (Лонд)
1988
336
176
179
[В паблике]
49
Андерсон
Г
,
Anderson
KL
,
Conroy
LA
,
Hallam
TJ
,
Moore
NC
,
Owen
JT
,
Jenkinson
EJ
Внутриклеточные сигнальные события при позитивной и негативной селекции CD4+CD8+тимоцитов in vitro0002
[В паблике]
50
Bendelac
A
,
Matzinger
P
,
Seder
RA
,
. тимусная селекция
J Exp Med
1992
175
731
742
[В паблике]
51
SWAT
W
,
DESSING
M
,
Von Boehmer
H
,
Kisielow
P
CD69 Arpression во время SELECTES и MATRATIONS 9000+
ARIMTERS
S
.
ARIMTERS
S
.
. Дж Иммунол
1993
23
739
746
[В паблике]
52
Brändle
D
,
Muller
S
,
Hengartner
H
,
Pircher
H
Regulation of RAG-1 and CD69 expression in the thymus during positive and negative selection
Eur J Immunol
1994
24
145
151
[В паблике]
53
Лебедь
KA
,
Альберола
J
-lla, J. A. Гросс, М. В. Эпплби, К.А. Форбуш, Дж.Ф. Томас и Р.М. Перлмуттер
Участие p21ras отличает положительную и отрицательную селекцию в тимоцитах
[В паблике]
54
Alberola-ILA
J
,
FORBUSH
KA
,
SEGER
R
,
Krebs
,
KREBS
,
KREBS 9000 2
,
Krebs 9000 2
,
,
.0002 RM
Селективная потребность в активации МАР-киназы при дифференцировке тимоцитов
[В паблике]
55
D’Ambrosio
D
,
Cantrell
DA
,
Frati
L
,
Santoni
A
,
Testi
R
Участие активации p21ras в экспрессии Т-клеточного CD69
Eur J Immunol
1994
24
616
620
[В паблике]
56
Chen
D
,
Rothenberg
EV
Molecular basis for developmental changes in interleukin-2 gene inducibility
Molec Cell Biol
1993
13
228
237
[В паблике]
57
Cosgrove
D
,
Chan
SH
,
Waltzinger
C
,
Benoist
C
,
Mathis
D
The thymic compartment ответственный за положительную селекцию CD4+T-клеток
Int Immunol
1993
4
707
710
[В паблике]
58
Marodon
G
,
Rocha
B
Generation of mature T cell populations in the thymus: CD4 or CD8 down-regulation occurs at different stages of thymocyte differentiation
Eur J Immunol
1994
24
196
204
[В паблике]
59
Пунт
JA
,
Осборн
BA
,
Такахама
Y
,
Sharrow
SO
,
Singer
A
Negative selection of CD4+CD8+thymocytes by T cell receptor-induced apoptosis requires a costimulatory signal that can be provided by CD28
J Exp Med
1994
179
709
713
[В паблике]
60
Surh
CD
,
Sprent
J
Т-клеточный апоптоз, обнаруженный в положении положительного и отрицательного отбора в тимусе
[В паблике]
61
Huang
R-P
,
Adamson
ED
Фосфорилированные формы фактора транскрипции Biophychem Egr-1 связываются с ДНК более эффективно, чем нефосфорилированные0003
1994
200
1271
1276
[В паблике]
62
Fehling
H J
,
Krotkova
A
,
Saint-Ruf
C
,
von Boehmer
H
Crucial role of the pre-T-cell ген рецептора альфа в развитии альфа-бета, но не гамма-дельта Т-клеток
Nature (Lond)
1995
375
795
798
[В паблике]
63
Skerka
C
,
Decker
EL
,
Zipfel
PF
Регуляторный элемент и промотчик для человека. EGR-1
J Biol Chem
1995
270
22500
22506
[В паблике]
64
Mages
HW
,
Stamminger
T
,
Rilke
O
,
Bravo
R
,
Kroczek
RA
Expression of PILOT , предполагаемый фактор транскрипции, требует двух сигналов и чувствителен к циклоспорину А в Т-клетках
[В паблике]
65
Lee
SL
,
Tourtellotte
LC
,
Wesselschmidt
RL
,
Milbrandt
J
Growth and differentiation proceeds normally in cells deficient in the immediate early ген NGFI-A
J Biol Chem
1995
270
9971
9977
[В паблике]
66
Шнайдер-Монури
S
,
Topilko
P
,
Seitanidou
T
,
Levi
G
,
Cohen-Tannoudji
T
,
Pournin
S
,
Babinet
C
,
Charnay
P
Нарушение Krox-20 приводит к изменению ромбомеров 3 и 5 в развивающемся
00030002 Сотовый
1993
75
1199
1214
[В паблике]
67
Kramer
B
,
Meichle
A
,
Hensel
G
,
Charnay
P
,
Kronke
M
Characterization of an Krox-24/Egr-1-чувствительный элемент в промоторе фактора некроза опухоли человека
Biochim Biophys Acta
1994
1219
413
421
[В паблике]
68
Maltzman
JS
,
Carmen
CA
,
Monroe
JG
Transcriptional regulation of the ICAM-1 gene in antigen receptor- and phorbol ester-stimulated B lymphocytes: role для фактора транскрипции EGR1
J Exp Med
1996
183
1747
1759
[В паблике]
69
Maltzman
JS
,
Carmen
CA
,
Monroe
JG
Role of EGR1 in regulation of stimulus-dependent CD44 transcription in B lymphocytes
Molec Cell Biol
1996
16
2283
2294
[В паблике]
70
Лопес-Кабрера
M
,
Munoz
E
,
Blazquez
MV
,
Ursa
MA
,
Santis
AG
,
Sanchez-Madrid
F
Транскрипционная регуляция гена, кодирующего человеческий лектиновый рецептор лейкоцитов С-типа AIM/CD69, и функциональная характеристика его элементов, чувствительных к фактору некроза опухоли-α
J Biol Chem
1995
270
21545
21551
[В паблике]
71
Williams
GT
,
LAU
LF
Активация индуцируемого гена рецептора орфан Nur77. 1993
13
6124
6136
[В паблике]
72
Calnan
BJ
,
Szychowski
S
,
Chan
FK-M
,
Cado
K
,
Winoto
A
A роль орфанного стероидного рецептора Nur77 в апоптозе, сопровождающем антиген-индуцированный негативный отбор
[В паблике]
73
CAO
x
,
Mahendran
R
,
Guy
Gr
,
Tan
YH
,
. сайт
J Biol Chem
1993
268
16949
16957
[В паблике]
74
МакМахон
СБ
,
Monroe
JG
Activation of the p21ras pathway couples antigen receptor stimulation to induction of the primary response gene egr-1 in B lymphocytes
J Exp Med
1995
181
417
422
[В паблике]
75
Gunter
KC
,
Irving
SG
,
Zipfel
PF
,
Siebenlist
0003
U
,
Kelly
K
Cyclosporin A-mediated inhibition of mitogeninduced gene transcription is specific for the mitogenic stimulus and cell type
J Immunol
1989
9
3286
3291
[В паблике]
76
Эммель
EA
,
Verweij
CL
,
Durand
3
DB
0003
Хиггинс
км
,
Lacy
E
,
Crabtree
GR
Циклоспорин A Определенно ингибирует функцию Nucelar Proteins, вовлеченные в T -клеточную активацию
Ncience Ncience Dcips).
246
1617
1620
[В паблике]
77
Вудроу
MA
,
Rayter
S
,
Downward
J
,
Cantrell
DA
p21ras function is important for T cell antigen receptor and protein kinase C regulation of nuclear factor of activated cells
J Immunol
1993
150
3853
3861
[В паблике]
78
Васкес
Нью-Джерси
,
Кай
Дж
,
Хедрик
SM
Клональная делеция двойных положительных тимоцитов in vivo и in vitro
[В паблике]
1 Сокращения, использованные в данной статье: CsA, циклоспорин А; Erg-1, реакция раннего роста.
Х. Шао и Д. Коно внесли равный вклад в эту работу.
Примечания автора
Адресная переписка с доктором Джонатаном Кеем, отделение иммунологии, IMM-8, Научно-исследовательский институт Скриппса, 10550 N. Torrey Pines Rd., La Jolla, California
.
данные и цифры
Данные и цифры
содержимое
Содержимое
добавки
Дополнения
ссылки
Ссылки
Рисунок 1
Ген Egr-1 быстро индуцируется после TCR-опосредованной активации двойной положительной клеточной линии DPK. ( A ) Тотальную РНК, выделенную из клеток DPK, культивируемых с иммобилизованным mAb против CD3ε в течение указанного времени (показанного в часах), подвергали анализу ОТ-ПЦР с использованием праймеров Egr-1 или CD4. ( B ) Анализ сдвига электрофоретической подвижности с использованием ядерных лизатов, полученных из клеток DPK, через 8 ч после активации иммобилизованным mAb против CD3ε. Зонды содержали один сайт связывания Egr-1 ( слева, ) или перекрывающиеся сайты Egr-1 и Sp1 ( справа, ). ( C ) Клетки DPK культивировали с клетками, представляющими фибробластный антиген DCEK-ICAM, и 1 мкМ голубиного цитохрома с пептида в течение указанного времени. Тотальную РНК выделяли и подвергали конкурентному анализу ОТ-ПЦР (см. Материалы и методы). Обратите внимание на разные шкалы экспрессии мРНК Egr-1 и CD4.
Рисунок 1
Ген Egr-1 быстро индуцируется после TCR-опосредованной активации двойной положительной клеточной линии DPK. ( A ) Тотальную РНК, выделенную из клеток DPK, культивируемых с иммобилизованным mAb против CD3ε в течение указанного времени (показанного в часах), подвергали анализу ОТ-ПЦР с использованием праймеров Egr-1 или CD4. ( B ) Анализ сдвига электрофоретической подвижности с использованием ядерных лизатов, полученных из клеток DPK, через 8 ч после активации иммобилизованным mAb против CD3ε. Зонды содержали один сайт связывания Egr-1 ( слева ) или перекрытие сайтов Egr-1 и Sp1 ( справа ). ( C ) Клетки DPK культивировали с клетками, представляющими фибробластный антиген DCEK-ICAM, и 1 мкМ голубиного цитохрома с пептида в течение указанного времени. Тотальную РНК выделяли и подвергали конкурентному анализу ОТ-ПЦР (см. Материалы и методы). Обратите внимание на разные шкалы экспрессии мРНК Egr-1 и CD4.
Закрыть модальное окно
Рисунок 2
Дифференцировка клеток DPK и 9Индукция мРНК 0363 Egr-2,3 чувствительна к циклоспорину А, тогда как индукция мРНК Egr-1 устойчива к циклоспорину А. ( A , B ) Клетки DPK культивировали с клетками, представляющими фибробластный антиген DCEK-ICAM, и 2 мкМ голубиного цитохрома с пептида в присутствии или в отсутствие 100 нг/мл циклоспорина А или подходящего разбавления растворителя (ДМСО) в качестве указано. Клетки собирали и окрашивали на CD69 через 1 или 3 дня культивирования ( A ) или окрашивали на CD4 и CD8 через 3 дня культивирования ().0363 В ). ( C ) ОТ-ПЦР-анализ тотальной РНК, полученной из клеток DPK, активированных в течение 6 часов иммобилизованным mAb против CD3ε в присутствии или в отсутствие 300 нг/мл циклоспорина А, с использованием Egr-1, Egr-2 (Krox- 20), праймеры CD4, CD69 или Egr-3. (*) Для указанных образцов также показан относительный уровень кДНК Egr-1, нормализованный к экспрессии кДНК CD4, как определено с помощью конкурентного анализа ОТ-ПЦР. Идентичность нижней основной полосы в ОТ-ПЦР Egr-3 подтверждали секвенированием.
Рисунок 2
Дифференцировка клеток DPK и Egr-2,3 Индукция мРНК чувствительна к циклоспорину А, в то время как индукция мРНК Egr-1 устойчива к циклоспорину А. ( A , B ) Клетки DPK культивировали с клетками, представляющими фибробластный антиген DCEK-ICAM, и 2 мкМ голубиного цитохрома с пептида в присутствии или в отсутствие 100 нг/мл циклоспорина А или подходящего разбавления растворителя (ДМСО) в качестве указано. Клетки собирали и окрашивали на CD69 через 1 или 3 дня культивирования ( A ) или окрашивали на CD4 и CD8 через 3 дня культивирования ( B ). ( C ) ОТ-ПЦР-анализ тотальной РНК, полученной из клеток DPK, активированных в течение 6 часов иммобилизованным mAb против CD3ε в присутствии или в отсутствие 300 нг/мл циклоспорина А, с использованием Egr-1, Egr-2 (Krox- 20), праймеры CD4, CD69 или Egr-3. (*) Для указанных образцов также показан относительный уровень кДНК Egr-1, нормализованный к экспрессии кДНК CD4, как определено с помощью конкурентного анализа ОТ-ПЦР. Идентичность нижней основной полосы в ОТ-ПЦР Egr-3 подтверждали секвенированием.
Закрыть модальное окно
Рисунок 3
Экспрессия доминантно-негативного мутанта p21ras блокирует дифференцировку клеток DPK. ( A ) RT-PCR анализ экспрессии N-ras в клетках DPK или тимоцитах, полученных от мышей дикого типа или мышей с дефицитом MHC. ( B ) Клеточные лизаты из клеток DPK и четырех независимых линий, которые экспрессируют Ha-ras N17, анализировали с помощью вестерн-блоттинга и зондировали антителами против ras. Эндогенный p21ras не виден на этом изображении. ( C ) Клетки DPK или 17N4 культивировали с клетками, презентирующими фибробластный антиген DCEK-ICAM, в присутствии ( жирные линии ) или в отсутствие ( тонкие линии ) 2 мкМ голубиного цитохрома с-пептида. Через 3 дня культивирования клетки собирали, окрашивали mAb к CD69 и анализировали с помощью проточной цитометрии. ( D ) Клетки DPK или 17N4 культивировали, как в ( C ), за исключением того, что клетки собирали на 1 или 3 день, как указано, и окрашивали mAb против CD4 и против CD8. Показаны проценты CD4 + 8 lo/- DPK клеток в обозначенных регионах.
Рисунок 3
Экспрессия доминантно-негативного мутанта p21ras блокирует дифференцировку клеток DPK. ( A ) RT-PCR анализ экспрессии N-ras в клетках DPK или тимоцитах, полученных от мышей дикого типа или мышей с дефицитом MHC. ( B ) Клеточные лизаты из клеток DPK и четырех независимых линий, которые экспрессируют Ha-ras N17, анализировали с помощью вестерн-блоттинга и зондировали антителами против ras. Эндогенный p21ras не виден на этом изображении. ( C ) Клетки DPK или 17N4 культивировали с клетками, презентирующими фибробластный антиген DCEK-ICAM, в присутствии ( жирные линии ) или в отсутствие ( тонкие линии ) 2 мкМ голубиного цитохрома с-пептида. Через 3 дня культивирования клетки собирали, окрашивали mAb к CD69 и анализировали с помощью проточной цитометрии. ( D ) Клетки DPK или 17N4 культивировали, как в ( C ), за исключением того, что клетки собирали на 1 или 3 день, как указано, и окрашивали mAb против CD4 и против CD8. Показаны проценты CD4 + 8 lo/- DPK клеток в обозначенных регионах.
Режим закрытия
Рисунок 4
Экспрессия семейства генов Egr зависит от сигнальных путей ras. ( A ) Конкурентную ОТ-ПЦР использовали для сравнения экспрессии генов Egr-1 и CD4 в клетках DPK или 17N4, активированных в течение 6 часов иммобилизованным mAb против CD3ε. ( B ) ОТ-ПЦР анализ тотальной РНК, полученной из клеток DPK или 17N4, активированных в течение 6 часов иммобилизованным mAb против CD3ε. Проводили независимые реакции ПЦР с использованием праймеров Egr-2 (Krox-20), CD4 или Egr-3.
Рисунок 4
Экспрессия семейства генов Egr зависит от сигнальных путей ras. ( A ) Конкурентную ОТ-ПЦР использовали для сравнения экспрессии генов Egr-1 и CD4 в клетках DPK или 17N4, активированных в течение 6 часов иммобилизованным mAb против CD3ε. ( B ) ОТ-ПЦР анализ тотальной РНК, полученной из клеток DPK или 17N4, активированных в течение 6 часов иммобилизованным mAb против CD3ε. Проводили независимые реакции ПЦР с использованием праймеров Egr-2 (Krox-20), CD4 или Egr-3.
Закрыть модальное окно
Рисунок 5
Активность связывания мРНК и ДНК Egr-1 в тимусе зависит от MHC. ( A ) Тимоциты, полученные от мышей дикого типа или мышей с дефицитом MHC, окрашивали в два цвета на CD4 и CD8. ( B ) Конкурентную ОТ-ПЦР использовали для определения уровня экспрессии генов CD4 и Egr-1 в тимоцитах, полученных от мышей дикого типа или мышей с дефицитом MHC. ( C ) Анализ сдвига электрофоретической подвижности с использованием ядерных лизатов, приготовленных из свежевыделенных тимоцитов, полученных от мышей дикого типа или мышей с дефицитом MHC, с использованием зонда, содержащего сайт связывания Egr-1. Ядерные экстракты, полученные из 5 × 10 9В реакциях связывания использовали клетки 0940 5 , содержащие эквивалентные количества белка. В некоторых случаях, как указано, реакции связывания содержали антитело против Egr-1 или нормальную кроличью сыворотку (NRS). Для сравнения показана реакция связывания, содержащая рекомбинантный Egr-1.
Рисунок 5
Активность связывания мРНК и ДНК Egr-1 в тимусе зависит от MHC. ( A ) Тимоциты, полученные от мышей дикого типа или мышей с дефицитом MHC, окрашивали в два цвета на CD4 и CD8. ( B ) Конкурентную ОТ-ПЦР использовали для определения уровня экспрессии генов CD4 и Egr-1 в тимоцитах, полученных от мышей дикого типа или мышей с дефицитом MHC. ( C ) Анализ сдвига электрофоретической подвижности с использованием ядерных лизатов, приготовленных из свежевыделенных тимоцитов, полученных от мышей дикого типа или мышей с дефицитом MHC, с использованием зонда, содержащего сайт связывания Egr-1. В реакциях связывания использовали ядерные экстракты, полученные из 5 × 10 5 клеток, содержащие эквивалентные количества белка. В некоторых случаях, как указано, реакции связывания содержали антитело против Egr-1 или нормальную кроличью сыворотку (NRS). Для сравнения показана реакция связывания, содержащая рекомбинантный Egr-1.
Закрыть модальное окно
Рисунок 6
Экспрессия мРНК Egr-1 в дважды положительных тимоцитах. ( A ) Общее количество тимоцитов и CD4 + 8 + тимоцитов (выделенных путем сортировки клеток, 95% DP), полученных от одного и того же животного, анализировали на экспрессию мРНК Egr и CD4 с помощью конкурентной ОТ-ПЦР. Показан относительный уровень кДНК Egr-1 в образце, нормализованный к уровню кДНК CD4. ( B ) Всего тимоцитов, полученных от мыши с дефицитом MHC, культивировали с гранулами, покрытыми хомячьим иммуноглобулином или анти-CD3ε mAb, в течение 90 минут до определения экспрессии генов Egr-1 и CD4 , как на рис. 1 C .
Рисунок 6
Экспрессия мРНК Egr-1 в дважды положительных тимоцитах. ( A ) Общее количество тимоцитов и CD4 + 8 + тимоцитов (выделенных путем сортировки клеток, 95% DP), полученных от одного и того же животного, анализировали на экспрессию мРНК Egr и CD4 с помощью конкурентной ОТ-ПЦР. Показан относительный уровень кДНК Egr-1 в образце, нормализованный к уровню кДНК CD4. ( B ) Всего тимоциты, полученные от мыши с дефицитом MHC, культивировали с гранулами, покрытыми хомячьим иммуноглобулином или анти-CD3ε mAb, в течение 90 минут перед определением экспрессии генов Egr-1 и CD4 , как показано на рис. 1 C .
Закрыть модальное окно
Рисунок 7
Экспрессия мРНК Egr-2 и Egr-3 в тимоцитах зависит от MHC. Тотальную РНК, полученную из свежевыделенных тимоцитов дикого типа или тимоцитов с нокаутом MHC, подвергали анализу ОТ-ПЦР с использованием праймеров CD4, Egr-2 (Krox-20) или Egr-3, как указано.
Рисунок 7
Экспрессия мРНК Egr-2 и Egr-3 в тимоцитах зависит от MHC. Тотальную РНК, полученную из свежевыделенных тимоцитов дикого типа или тимоцитов с нокаутом MHC, подвергали анализу ОТ-ПЦР с использованием праймеров CD4, Egr-2 (Krox-20) или Egr-3, как указано.
Закрыть модальное окно
Рисунок 8
Экспрессия белка Egr-1 в тимусе. Тонкие срезы нормального тимуса ( A–C ) или тимуса с нокаутом MHC ( D ) фиксировали в формальдегиде и окрашивали антисывороткой к специфическому кроличьему пептиду против Egr-1 ( A , C , D ) или тем же антителом, предварительно инкубированным со специфичным пептидом ( B ). Обозначены области коры ( C ) и мозгового вещества ( M ). Срезы контрастно окрашивали гематоксилином и фотографировали с увеличением ×20 ( A , B ) или ×40 ( C , D ). C показывает увеличение того же участка, сфотографированного в А .
Рисунок 8
Экспрессия белка Egr-1 в тимусе. Тонкие срезы нормального тимуса ( A–C ) или тимуса с нокаутом MHC ( D ) фиксировали в формальдегиде и окрашивали специфичной кроличьей антисывороткой против пептида Egr-1 ( A , C , D ). ) или то же самое антитело, предварительно инкубированное со специфическим пептидом ( B ). Обозначены области коры ( C ) и мозгового вещества ( M ). Срезы докрашивали гематоксилином и фотографировали с увеличением ×20 ( A , B ) или ×40 ( C , D ). C показывает увеличенное изображение той же секции, сфотографированной в A .
Закрыть модальное окно
Рисунок 9
Экспрессия белка Egr-1 в субпопуляциях тимоцитов. Тимоциты молодой взрослой мыши окрашивали в 4 цвета для выявления экспрессии CD4, CD8, Egr-1 и CD69 или CD3 и анализировали с помощью FACS ®, как описано в разделе «Материалы и методы». На точечных графиках указано процентное содержание тимоцитов в каждом квадранте или в скобках ( верхний правый точечный график ), процент Egr-1 + тимоцитов в каждой подгруппе тимоцитов. Там, где указано, окрашивание показано для гейтированных популяций тимоцитов (либо Egr-1 + тимоцитов, как показано на гистограмме, либо CD4 + 8 + тимоцитов). Аналогичные результаты были получены от трех других отдельных животных.
Рисунок 9
Экспрессия белка Egr-1 в субпопуляциях тимоцитов. Тимоциты молодой взрослой мыши окрашивали в 4 цвета для выявления экспрессии CD4, CD8, Egr-1 и CD69. или CD3 и анализировали с помощью FACS ® , как описано в разделе «Материалы и методы». На точечных графиках указан процент тимоцитов в каждом квадранте или в скобках ( вверху справа точечный график ) процент Egr-1 + тимоцитов в каждом поднаборе тимоцитов. Там, где указано, окрашивание показано для гейтированных популяций тимоцитов (либо Egr-1 + тимоцитов, как показано на гистограмме, либо CD4 + 8 + тимоцитов). Аналогичные результаты были получены от трех других отдельных животных.
Закрыть модальное окно
- Предыдущая статья
- Следующая статья
1
Sukhatme
VP
,
Kartha
S
,
Toback
FG
,
Taub
R
,
Hoover
RG
,
Tsai-Morris
C-H
A novel early growth response gene rapidly induced by fibroblast, epithelial cell and lymphocyte mitogens
Oncogene Res
1987
1
343
355
[В паблике]
2
Sukhatme
VP
,
CAO
x
,
Chang
LC
,
TSAI-Morris
0003
C-H
,
Stamenkovich
D
,
Ferreira
PCP
,
Cohen
DR
,
Edwards
SA
,
Shows
TB
,
Curran
T
et al
Ген, кодирующий цинковые пальцы, который корегулируется c-fos во время роста и дифференцировки и после клеточной деполяризации
Клетка
1988
53
37
43
[В паблике]
3
Christy
B
,
Nathans
LF
,
Nathans
D
Активированные мыши 3T3 Факторы роста. Proc Natl Acad Sci USA
1988
85
7857
7861
[В паблике]
4
Lemaire
P
,
Откровение
O
,
Bravo
R
,
Charnay
P
,
Charnay
P
,
Charnay
P
,
Charnay
P
,
,
,
. домены активируются факторами роста в культивируемых клетках
Proc Natl Acad Sci USA
1988
85
4691
4695
[В паблике]
5
Lim
RW
,
Varnum
BC
,
Herschman
HR
Cloning of tetradecanoyl phorbol ester-induced ‘primary response’ sequences and their expression in density-arrested Swiss 3T3 клетки и непролиферативный вариант ТРА
Онкоген
1987
1
263
270
[В паблике]
6
Милбрандт
J
Ген, индуцированный фактором роста нервов, кодирует возможный фактор транскрипции
[В паблике]
7
Irving
SG
,
June
CH
,
Zipfel
PF
,
Siebenlist
U
,
Kelly
K
Гены, индуцированные митогеном, подвергаются многочисленным путям регуляции на начальных стадиях активации Т-клеток
[В паблике]
8
Cao
X
,
Koski
RA
,
Gashler
A
,
McKiernan
M
,
Morris
CF
,
Gaffney
R
,
Hays
RV
,
SUKHATME
VP
Идентификация и характеристика Gene-BIND-BINDININGE-BINDININGE-BINDININGE-BINDININGE-BINDIND-BINDIND-BINDIND-BINDININGE-BINDININGE-BINDIND-BINDIND-BINDINDINGE-BINDININ индуцируется сигналами дифференцировки и роста
Mol Cell Biol
1990
10
1931
1939
[В паблике]
9
Кристи
B
,
Nathans
D
DNA binding site of the growth factor-inducible protein Zif268
Proc Natl Acad Sci USA
1989
86
8737
8741
[В паблике]
10
Павлетич
NP
,
Pabo
CO
Цинковый палец-ДНК распознавание: кристаллическая структура комплекса Zif26W 9 DC0029 при 2,03 A Science (0003
1991
252
809
817
[В паблике]
11
Swirnoff
AH
,
Milbrandt
J
DNA-binding specificity of NGFI-A and related zinc-finger transcription factors
Mol Cell Biol
1995
15
2275
2287
[В паблике]
12
Гашлер
AL
,
Swaminathan
S
,
Sukhatme
VP
Новый модуль репрессии, обширный домен активации и двудольный сигнал ядерной локализации, определенный в прототипе E-early GR1 транскрипции: Семейство цинковых факторов транскрипции
Cell Biol
1993
13
4556
4571
13
Russo
МВт
,
0003
Matheny
C
,
Milbrandt
J
Transcriptional activity of the zinc finger NGFI-A is influenced by its interaction with a cellular factor
Molec Cell Biol
1993
13
6858
6865
[В паблике]
14
Carman
JA
,
Monroe
JG
Белок EGR1 содержит дискретный транскрипционный регуляторный домен, делеция которого приводит к укороченному белку, который блокирует транскрипцию9 EGR10003
DNA Cell Biol
1995
14
581
589
[В паблике]
15
Russo
MW
,
Sevetson
BR
,
Milbrandt
J
Идентификация NAB1, Repressor of NGFI-A-ANDIC202. Наука США
1995
92
6873
6877
[В паблике]
16
Chavrier
P
,
Zerial
M
,
Lemaire
P
,
Almendral
J
,
Bravo
R
,
Charnay
P
Ген, кодирующий белок с цинковыми пальцами, активируется при переходе G0/G1 в культивируемых клетках
EMBO (Eur Mol Biol Organ)
1988
7
29
35
[В паблике]
17
Chavrier
P
,
Janssen-Timmen
U
,
Mattei
M-G
,
Zerial
M
,
Bravo
R
,
Charnay
P
Структура, расположение хромосом и экспрессия гена цинковых пальцев мыши Krox20: множественные генные продукты и корегуляция с протоонкогеном c-fos
Мол Селл Биол
1989
9
787
797
[В паблике]
18
Joseph
LJ
,
Le Beau
MM
,
Jamieson
GA
Jr,
Acharya
S
,
Shows
TB
,
Роули
JD
,
Сукхатме
VP
Молекулярное клонирование, секвенирование и картирование EGR2, гена реагирования на ранний рост человека, кодируя белок с структурой «цинкерующий пальцы»
Proc Natl Acad Sci USA
1988
85
7164
7168
7164
7168
7164
7168
9000 2
7164
7168
9000 2
[В паблике]
19
Zipfel
PF
,
Irving
SG
,
Kelly
K
,
SiebenList
,
Siebenlist
9000
,
Siebenlist
9000 9000
,
Siebenlist
9000 2
,
. 0002 U
Сложность первичного генетического ответа на митогенную активацию Т-клеток человека
Mol Cell Biol
1989
9
1041
1048
[В паблике]
20
Patwardhan
S
,
Gashler
A
,
Siegel
MG
,
Chang
LC
,
Joseph
LJ
,
показывает
TB
,
Le Beau
MM
,
Sukhatme
VP
EGR3, новое число семейства EGRENERENTIONSINGINGINGINGINGINGINGINGINGENTIONNICTION
.
1991
6
917
928
[В паблике]
21
Кросби
SD
,
Путц
JJ
,
Simburger
KS
,
Fahrner
TJ
,
Milbrandt
J
The early response gene NGFI-C encodes a zinc finger transcriptional activator and is a member of the GCGGGGGCG (GSG ) семейство белков, связывающих элементы
Mol Cell Biol
1991
11
3835
3841
[В паблике]
22
Мюллер
H-J
,
Skerka
C
,
Bialonski
A
,
Zipfel
PF
Clone PAT 133 Идентифицируют ген Gene, который Engodes Other Cumber-Clase-Clone-Clone-Gene-Factore-Clase-Clase-Factore-Inmeded-Clectude-Clase-Factore-Clase-Factore-Clase-Factore-Clase-Factore-Clase. почти идентичные домены цинковых пальцев
Proc Natl Acad Sci USA
1991
88
10079
10083
[В паблике]
23
Гашлер
A
,
SUKHATME
VP
Ранний белок ответа на ранний рост 1 (EGR-1): прототип семейства цинковых фантазий. Факторы транскрипции
.
224
[В паблике]
24
NGUYEN
HQ
,
Hoffman-Liebermann
B
,
Liebermann
DA
Транскрипционный фактор Zinc Film0003
Сотовый
1993
72
197
209
[В паблике]
25
Krishnaraju
K
,
Nguyen
HQ
,
Liebermann
DA
,
Hoffman
DA
,
DA
,
DA
,
DA
,
DA
,
9000. гемопоэтические клетки
Molec Cell Biol
1995
15
5499
5507
[В паблике]
26
Cui
M-Z
,
Parry
GCN
,
Oeth
P
,
Larson
H
,
Smith
M
,
Хуан
Р-П
,
Адамсон
ЭД
,
Макман
N
Транскрипционная регуляция гена тканевого фактора в эпителиальных клетках человека опосредована Sp1 и EGR-1
[В паблике]
27
KHACHIGIAN
LM
,
Lindner
V
,
Williams
AJ
,
Collins
T.
,
Collins
T.
,
. при повреждении сосудов
Наука (Ваш, округ Колумбия)
1996
271
1427
1431
[В паблике]
28
Sukhatme
VP
Ранние события транскрипции в росте клеток: семейство EGR
J Am Soc Nephrol
1990
1
859
866
[В паблике]
29
Сейферт
ВЛ
,
Сукхатме
VP
,
Monroe
JG
Differential expression of a zinc finger-encoding gene in response to positive versus negative signaling through receptor imunoglobulin in murine B lymphocytes
Molec Cell Biol
1989
9
2083
2088
[В паблике]
30
Seyfert
VL
,
McMahon
S
,
GLENN
W
,
CAO
X
,
SUKHATME
VP
,
Monroe
J
EGR-MERVER-Activation in Surface гро с активацией протеинкиназы С
J Иммунол
1990
145
3647
3653
[В паблике]
31
Маковер
Д
,
Cuddy
M
,
Yum
S
,
Bradley
K
,
Alpers
J
,
Sukhatme
V
,
Reed
JC
Опосредованное эфиром форбола ингибирование роста и регуляция экспрессии протоонкогена в линии Т-клеточного лейкоза человека JURKAT
Oncogene
1991
6
455
460
[В паблике]
32
Perez-Castillo
AP
,
Pipaon
C
,
Garcia
I
,
Alemany
I
,
Alemany
Im. пролиферация
J Biol Chem
1993
268
19445
19450
[В паблике]
33
Seyfert
VL
,
McMahon
SB
,
Glenn
WD
,
Yellen
AJ
,
Sukhatme
VP
,
Cao
XM
,
Monroe
JG
Метилирование немедленно-раннего индуцируемого гена как механизм индукции толерантности В-клеток
Science (Wash DC)
1990
250
797
800
[В паблике]
34
Klaus
SJ
,
Phillips
NE
,
Parker
DC
Эффекты IL-4 и FC GAMMA Antagement II Antagement на EGR-1. методом кросслинкинга мембранного иммуноглобулина
Molec Immunol
1993
30
1553
1558
[В паблике]
35
GOTSCHALK
AR
,
JOSEPH
LJ
,
Quintans
J
FCGRII поперечный сцепление ингибирует ингибирует ANDI-IG-индуцированные EGR-1 EGR-1. Дж Иммунол
1994
152
2115
2122
[В паблике]
36
Дженкинс
МК
,
Schwartz
RH
,
Pardoll
DM
Effects of cyclosporin A on T-cell development and clonal deletion
Science (Wash DC)
1988
241
1655
1658
[В паблике]
37
Watson
MA
,
Milbrandt
J
Экспрессия генов NGFI-A и NGFI-B, регулируемых фактором роста нервов9, у развивающихся крыс
Развитие
1990
110
173
183
[В паблике]
38
Kaye
J
,
Ellenberger
DE
Differentiation of an immature T cell line: a model of thymic positive selection
Cell
1992
71
423
435
[В паблике]
39
Грусби
MJ
,
Johnson
RS
,
Papaioannou
VE
,
Glimcher
LH
Истощение CD4+T-клетки. Мойка DC)
1991
253
1417
1420
[В паблике]
40
Зейлстра
М
,
Бикс
M
,
Simister
NE
,
LORING
JM
,
RAULET
DH
,
JAENISCH 9000 3 9000.103INRELG2 9000. 9000. 9000. 9000. 9000. 9000. 9000. 9000. 9000.
,
JAENISCH 9000 3 9000 2
,
JAENISCH 9000 9000.
,
. цитолитические Т-клетки
Природа (Лонд)
1990
344
742
746
[В паблике]
41
ДеКонинг
Дж
,
Carbone
F
,
Kaye
J
Контраст между MHC-опосредованной дифференцировкой CD4+CD8+T-клеточной линии класса I и класса II: последствия для клональной приверженности
Int
7
541
549
[В паблике]
42
Quilliam
LA
,
Като
K
,
Рабун
км
,
HISAKA
мм
,
HUFF
SY
,
Campbell-Burk
S
,
Der
CJ
,
S
,
,
,
,
. ) ингибирующий рост фенотип и взаимодействие ras с факторами обмена гуаниновых нуклеотидов
[В паблике]
43
Kuhlman
P
,
Moy
VT
,
Lollo
BA
,
Brian
AA
The accessory function of murine intercellular adhesion molecule-1 in T активация лимфоцитов
Дж Иммунол
1991
146
1773
1782
[В паблике]
44
Хубэнк
М
,
Schatz
DG
Идентификация различий в экспрессии мРНК с помощью репрезентативного разности анализа CDNA
NUC ACIDS RES
1994
22
5640
5649
[В паблике]
45
Kaye
J
,
HSU
M-L
,
SAURON
M-E
,
SC
,
,
,
,
,
,
,
,
. 0003
Gascoigne
NRJ
,
Hedrick
SM
Selective development of CD4+T cells in transgenic mice expressing a class II MHC-restricted antigen receptor
Nature (Lond)
1989
341
746
749
[В паблике]
46
Пуарье
G
,
Lo
D
,
Рейли
CR0003
,
Kaye
J
Discrimination between thymic epithelial cells and peripheral antigen presenting cells in the induction of immature T cell differentiation
Immunity
1994
1
385
391
[В паблике]
47
ДеКонинг Дж. и Дж. Кэй. Требования к дифференцировке незрелой линии CD4+8+ Т-клеток. Дев. Иммунол. Под давлением.
48
GAO
EK
,
LO
D
,
CHENEY
R
,
SHANAGAWA
,
Shrest.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
9000.
.
. с циклоспорином А
Nature (Лонд)
1988
336
176
179
[В паблике]
49
Андерсон
Г
,
Anderson
KL
,
Conroy
LA
,
Hallam
TJ
,
Moore
NC
,
Owen
JT
,
Jenkinson
EJ
Внутриклеточные сигнальные события при позитивной и негативной селекции CD4+CD8+тимоцитов in vitro
J Иммунол
1995
154
3636
3643
[В паблике]
50
Bendelac
A
,
Matzinger
P
,
Seder
RA
,
. тимусная селекция
J Exp Med
1992
175
731
742
[В паблике]
51
SWAT
W
,
DESSING
M
,
VON BOEHMER
H
,
KISIELOW
P
,
KISIELOW
P
. тимоциты
Eur J Immunol
1993
23
739
746
[В паблике]
52
Брендле
D
,
Muller
S
,
Hengartner
H
,
Pircher
H
Regulation of RAG-1 and CD69 expression in the thymus during positive and negative selection
Eur J Immunol
1994
24
145
151
[В паблике]
53
Лебедь
KA
,
Альберола
J
-lla, J. A. Гросс, М. В. Эпплби, К.А. Форбуш, Дж.Ф. Томас и Р.М. Перлмуттер
Участие p21ras отличает положительную и отрицательную селекцию в тимоцитах
[В паблике]
54
Alberola-ILA
J
,
FORBUSH
KA
,
SEGER
R
,
Krebs
,
KREBS
,
KREBS 9000 2
,
Krebs 9000 2
,
,
.0002 RM
Селективная потребность в активации МАР-киназы при дифференцировке тимоцитов
[В паблике]
55
D’Ambrosio
D
,
Cantrell
DA
,
Frati
L
,
Santoni
A
,
Testi
R
Участие активации p21ras в экспрессии Т-клеточного CD69
Eur J Immunol
1994
24
616
620
[В паблике]
56
Chen
D
,
Rothenberg
EV
Molecular basis for developmental changes in interleukin-2 gene inducibility
Molec Cell Biol
1993
13
228
237
[В паблике]
57
Cosgrove
D
,
Chan
SH
,
Waltzinger
C
,
Benoist
C
,
Mathis
D
The thymic compartment ответственный за положительную селекцию CD4+T-клеток
Int Immunol
1993
4
707
710
[В паблике]
58
Marodon
G
,
Rocha
B
Generation of mature T cell populations in the thymus: CD4 or CD8 down-regulation occurs at different stages of thymocyte differentiation
Eur J Immunol
1994
24
196
204
[В паблике]
59
Пунт
JA
,
Осборн
BA
,
Такахама
Y
,
Sharrow
SO
,
Singer
A
Negative selection of CD4+CD8+thymocytes by T cell receptor-induced apoptosis requires a costimulatory signal that can be provided by CD28
J Exp Med
1994
179
709
713
[В паблике]
60
Surh
CD
,
Sprent
J
Т-клеточный апоптоз, обнаруженный в положении положительного и отрицательного отбора в тимусе
[В паблике]
61
Huang
R-P
,
Adamson
ED
Фосфорилированные формы фактора транскрипции Biophychem Egr-1 связываются с ДНК более эффективно, чем нефосфорилированные0003
1994
200
1271
1276
[В паблике]
62
Fehling
H J
,
Krotkova
A
,
Saint-Ruf
C
,
von Boehmer
H
Crucial role of the pre-T-cell ген рецептора альфа в развитии альфа-бета, но не гамма-дельта Т-клеток
Nature (Lond)
1995
375
795
798
[В паблике]
63
Skerka
C
,
Decker
EL
,
Zipfel
PF
Регуляторный элемент и промотчик для человека. EGR-1
J Biol Chem
1995
270
22500
22506
[В паблике]
64
Mages
HW
,
Stamminger
T
,
Rilke
O
,
Bravo
R
,
Kroczek
RA
Expression of PILOT , предполагаемый фактор транскрипции, требует двух сигналов и чувствителен к циклоспорину А в Т-клетках
[В паблике]
65
Lee
SL
,
Tourtellotte
LC
,
Wesselschmidt
RL
,
Milbrandt
J
Growth and differentiation proceeds normally in cells deficient in the immediate early ген NGFI-A
J Biol Chem
1995
270
9971
9977
[В паблике]
66
Шнайдер-Монури
S
,
Topilko
P
,
Seitanidou
T
,
Levi
G
,
Cohen-Tannoudji
T
,
Pournin
S
,
Babinet
C
,
Charnay
P
Нарушение Krox-20 приводит к изменению ромбомеров 3 и 5 в развивающемся
00030002 Сотовый
1993
75
1199
1214
[В паблике]
67
Kramer
B
,
Meichle
A
,
Hensel
G
,
Charnay
P
,
Kronke
M
Characterization of an Krox-24/Egr-1-чувствительный элемент в промоторе фактора некроза опухоли человека
Biochim Biophys Acta
1994
1219
413
421
[В паблике]
68
Maltzman
JS
,
Carmen
CA
,
Monroe
JG
Transcriptional regulation of the ICAM-1 gene in antigen receptor- and phorbol ester-stimulated B lymphocytes: role для фактора транскрипции EGR1
J Exp Med
1996
183
1747
1759
[В паблике]
69
Maltzman
JS
,
Carmen
CA
,
Monroe
JG
Role of EGR1 in regulation of stimulus-dependent CD44 transcription in B lymphocytes
Molec Cell Biol
1996
16
2283
2294
[В паблике]
70
Лопес-Кабрера
M
,
Munoz
E
,
Blazquez
MV
,
Ursa
MA
,
Santis
AG
,
Sanchez-Madrid
F
Транскрипционная регуляция гена, кодирующего человеческий лектиновый рецептор лейкоцитов С-типа AIM/CD69, и функциональная характеристика его элементов, чувствительных к фактору некроза опухоли-α
J Biol Chem
1995
270
21545
21551
[В паблике]
71
Williams
GT
,
LAU
LF
Активация индуцируемого гена рецептора орфан Nur77. 1993
13
6124
6136
[В паблике]
72
Calnan
BJ
,
Szychowski
S
,
Chan
FK-M
,
Cado
K
,
Winoto
A
A роль орфанного стероидного рецептора Nur77 в апоптозе, сопровождающем антиген-индуцированный негативный отбор
[В паблике]
73
CAO
x
,
Mahendran
R
,
Guy
Gr
,
Tan
YH
,
. сайт
J Biol Chem
1993
268
16949
16957
[В паблике]
74
МакМахон
СБ
,
Monroe
JG
Activation of the p21ras pathway couples antigen receptor stimulation to induction of the primary response gene egr-1 in B lymphocytes
J Exp Med
1995
181
417
422
[В паблике]
75
Gunter
KC
,
Irving
SG
,
Zipfel
PF
,
Siebenlist
0003
U
,
Kelly
K
Cyclosporin A-mediated inhibition of mitogeninduced gene transcription is specific for the mitogenic stimulus and cell type
J Immunol
1989
9
3286
3291
[В паблике]
76
Эммель
EA
,
Verweij
CL
,
Durand
3
DB
0003
Хиггинс
км
,
Lacy
E
,
Crabtree
GR
Циклоспорин A Определенно ингибирует функцию Nucelar Proteins, вовлеченные в T -клеточную активацию
Ncience Ncience Dcips).
246
1617
1620
[В паблике]
77
Вудроу
MA
,
Rayter
S
,
Downward
J
,
Cantrell
DA
p21ras function is important for T cell antigen receptor and protein kinase C regulation of nuclear factor of activated cells
J Immunol
1993
150
3853
3861
[В паблике]
78
Васкес
Нью-Джерси
,
Кай
Дж
,
Хедрик
SM
Клональная делеция двойных положительных тимоцитов in vivo и in vitro
[В паблике]
EGR 8 Анализ: Hoosiers by the Numbers
EGR 8 состоит из округов Браун, Дэвисс, Грин, Лоуренс, Мартин, Монро, Ориндж и Оуэн.
EGR 8 расположен в живописном южно-центральном районе Индианы, посреди холмистой местности и красивых природных зон.
Этот регион исторически был центром добычи известняка в Соединенных Штатах, с карьерами в Блумингтоне, Бедфорде и Эллетсвилле, и это лишь некоторые из них. Промышленность была настолько важна для региона, что стала известна как «Каменный пояс», который простирается на север до округа Патнэм и на юг через округа Оуэн, Монро, Лоуренс и Ориндж. Некоторые из самых известных построек из известняка в мире были построены с использованием известняка Индианы, в том числе Национальный собор, Эмпайр-стейт-билдинг, Пентагон и 35 из 50 зданий Капитолия штата в США.0003
Сегодня промышленная база региона включает образование, здравоохранение, производство и туризм. Во флагманском кампусе Университета Индианы, IU-Bloomington, обучается более 40 000 студентов, и он является крупнейшим работодателем в регионе. Индустрия туризма также имеет большое значение: исторический курорт French Lick Springs Resort и отель West Baden Springs, а также всемирно известные поля для гольфа, созданные Питом Даем и Дональдом Россом, расположены в округе Ориндж.
Другими крупными работодателями в регионе являются Cook Group, IU Health Bloomington Hospital, Boston Scientific Corporation и NAVSEA Crane. Учебными заведениями региона являются IU Bloomington и Ivy Tech.
Недавний анализ
Рынок труда
Отзыв
июль 2022 г.
Снимок сектора промышленности
Четвертый квартал 2021 года
Статистика профессиональной занятости
2021 Годовой
Доступ к архивным публикациям с помощью Инструмента поиска публикаций.
Быстрая статистика
Щелкните любую краткую статистику, чтобы просмотреть подробности и сравнительную таблицу.
315 736
Население (ЕГР 8)
2,9
Уровень безработицы (EGR 8)
$15. 0
млрд Личный доход (EGR 8)
25 долларов США
Средняя заработная плата (Блумингтон MSA)
$18,63
Средняя заработная плата (Блумингтон MSA)
78 200
Nonfarm Employment (Bloomington MSA)
Свяжитесь с региональным аналитиком
Нужна помощь? Знающие региональные аналитики могут предложить помощь.
Кэсси Джейнс
Электронная почта: [email protected]
Телефон: (765) 507-9710
Электронная почта Кэсси
Основные полученные гранты
- Расширение JAG – 437 926 долларов США
- Грант Workforce Ready (WRG)
– 434 000 долларов - Грант на поддержку производительности
– 485 284 - NLJ — Грант на обучение работодателя
– 970 000 долларов США
долларов США
10 ведущих профессиональных групп по росту
(долгосрочные прогнозы на 2020-2030 гг. )
- Личный уход и обслуживание
- Профессии, связанные с оказанием медицинской помощи
- Профессии, связанные с приготовлением пищи и обслуживанием
- Искусство, дизайн, развлечения, спорт и СМИ
- Занятия в сфере общественных и социальных услуг
- Компьютерные и математические занятия
- Занятия по уборке и обслуживанию зданий и территорий
- Юридические занятия
- Медицинские работники и технические специалисты
- Образование, обучение и библиотечные профессии
Источник: Бюро статистики труда США и Департамент развития рабочей силы Индианы
Узнайте о долгосрочных прогнозах занятости с помощью Инструмента прогнозирования перспектив занятости .
10 ведущих отраслей промышленности
(На основе среднегодовой занятости в 2021 г. )
- Производство
- Здравоохранение и социальные услуги
- Розничная торговля
- Услуги по размещению и питанию
- Государственное управление
- Строительство
- Профессиональный, научный, технический
- Образовательные услуги
- Прочие услуги (кроме государственного управления)
- Транспорт и складское хозяйство
Источник: Ежеквартальная перепись занятости и заработной платы
Доступ к нескольким наборам данных на отраслевой тематической странице .
Крупные работодатели
(на основе количества сотрудников)
Найдите еще больше работодателей по местонахождению, классу размера или отрасли с помощью инструмента Business Lookup Tool .
EGR 8
- Университет Индианы Блумингтон
- Кук Групп Инк.
- Каталант
- Больница IU Health Блумингтон
- French Lick Spring Resort & Spa
- Вест Баден Спрингс Отель
- Бостонская научная корпорация
- Трансмиссия GM
- Кран NAVSEA
- Бакстер Биофарма Солюшнс
О данных
Примечание. Список региональных работодателей на этой странице был обновлен в июле 2022 г., чтобы включить местные данные.
Эта база данных содержит списки почти 12 миллионов работодателей США. Информация о работодателе предоставлена компанией Data Axle ® , Омаха, Небраска, (866)-DATAXLE. Copyright © 2022 . Все права защищены.
Основные учебные заведения
Наименование | Тип | Город |
|---|---|---|
| Университет Индианы Блумингтон | Четырехлетний | Блумингтон |
| Ivy Tech — Блумингтон | Двухлетний | Блумингтон |
Источник: Комиссия по высшему образованию Индианы
Используйте
INtraining для поиска
программы обучения и обучения
Крупнейшие города в регионе экономического роста 8
| Имя | Население в 2021 году | процентов региона | округ |
|---|---|---|---|
| Блумингтон | 79 968 | 25,3% | Монро |
| Бедфорд | 13 794 | 4,4% | Лоуренс |
| Вашингтон | 12 040 | 3,8% | Дэвис |
| Эллетсвилл | 6 588 | 2,1% | Монро |
| Линтон | 5 147 | 1,6% | Грин |
| Митчелл | 3 934 | 1,2% | Лоуренс |
| Паоли | 3625 | 1,1% | Оранжевый |
| Лугути | 2 599 | 0,8% | Мартин |
| Спенсер | 2 426 | 0,8% | Оуэн |
| Блумфилд | 2 251 | 0,7% | Грин |
Источник: Ежегодные оценки численности населения Бюро переписи населения США
Courthouse Square, Bloomington, Indiana
Полезные региональные инструменты и ресурсы сила, жилье и многое другое.
Основные сведения о округе
Получите моментальный снимок критической статистики населения и рабочей силы для каждого округа в регионе.
- Коричневый
- Дэвис
- Грин
- Лоуренс
- Мартин
- Монро
- Оранжевый
- Оуэн
Indiana Career Ready INDemand Jobs
Найдите объявления о вакансиях и тренинги для высокооплачиваемых и востребованных вакансий.
Региональный состав рабочей силы
Узнайте региональный состав профессий и/или отраслей, чтобы получить представление о сильных сторонах или выявить пробелы.
Regionizer
Создайте регион для просмотра демографических данных, моделей поездок на работу, отраслевых кластеров и кластеров занятий, включая возможность включения округов, граничащих с Индианой.
Прогнозы рабочей силы
Это официальные прогнозы рабочей силы до 2050 года, опубликованные в ноябре 2018 года Центром бизнес-исследований Индианы.
Коммутация
Страница данных о поездках обеспечивает легкий доступ к множеству наборов данных о поездках и визуализациях.
Региональные ссылки
Просмотреть руководство от А до Я для списка внешних ссылок по всему штату.
- Вестник Таймс
География | Ежечасно | Годовой |
|---|---|---|
| Блумингтон MSA | 25,00 $ | 52 000 долларов |
| Индиана | 19,05 $ | $50 143 |
| США | 28,01 $ | $58 260 |
Источник: OES от Бюро статистики труда США и Департамента развития рабочей силы штата Индиана
Географическое положение | |
|---|---|
| Блумингтон MSA | 78 200 |
| Индиана | 3 194 600 |
| США | 152 572 000 |
Примечание: Данные без сезонной корректировки
Источник: Бюро статистики труда США и Департамент развития рабочей силы Индианы
Географическое положение | |
|---|---|
| Район 8 | $15 027,0 |
| Индиана | $350 759,0 |
| США | $19 607 447,0 |
Источник: Бюро экономического анализа США
Географическое положение | |
|---|---|
| Район 8 | 315 736 |
| Индиана | 6 805 985 |
| США | 331 893 745 |
Источник: Бюро переписи населения США
Географическое положение | Ежечасно | Годовой |
|---|---|---|
| Блумингтон MSA | $18,63 | $38 750 |
| Индиана | $15,23 | 45 449 долларов |
| США | 22,00 $ | 45 760 долларов |
Источник: OES от Бюро статистики труда США и Департамента развития рабочей силы штата Индиана
Географическое положение | |
|---|---|
| Район 8 | 2,9 |
| Индиана | 3,1 |
| США | 3,8 |
Примечание: Данные без сезонной корректировки
Источник: Бюро статистики труда США и Департамент развития рабочей силы Индианы
Использование одностороннего клапана для оптимизации производительности турбонагнетателя на двигателе со смешанной системой рециркуляции отработавших газов
Введение
Все более строгие нормы выбросов в дизельных двигателях привели к исследованиям в области технологии рециркуляции отработавших газов (EGR) для уменьшения выхода двигателя из строя NO X . Термическое, химическое и разбавляющее воздействие рециркуляции отработавших газов помогает уменьшить выброс NO X в двигателе. Традиционно в этом приложении на протяжении нескольких десятилетий использовалась система рециркуляции отработавших газов высокого давления (Park et al., 2014). Различные компоновки EGR были опробованы для проверки их влияния на производительность и выбросы двигателя (Mao et al., 2015; Zheng et al., 2004; Vitek et al., 2008) и концепция двойного контура, которая включает в себя высокую Из них рождается контур EGR с давлением (HP-EGR) и контур EGR с низким давлением (LP-EGR).
В системе LP-EGR выхлопные газы удаляются со стороны низкого давления турбины и подаются на сторону низкого давления компрессора (рис. 1А). Основное преимущество системы LP-EGR заключается в том, что турбина согласована, и, следовательно, перепад давления в двигателе не изменяется. Поскольку размер компрессора может отличаться от размера турбины, система LP-EGR может обеспечить большое количество EGR, особенно при низких скоростях и условиях высокой нагрузки (Williams et al. , 2017). Кроме того, поскольку линия подачи LP-EGR длиннее, чем у HP-EGR, смешивание газа LP-EGR и свежего воздуха улучшается, что приводит к более однородной смеси в цилиндрах. По сравнению с системой HP-EGR, система LP-EGR также оказывает меньшее влияние на энтальпию на входе в турбину, поскольку скорость EGR варьируется. Таким образом, изменение скорости вращения турбонагнетателя, эффективности компрессора и PMEP минимально по мере увеличения скорости EGR, и, как правило, может быть достигнуто более высокое давление наддува (Mao et al., 2015; Maiboom et al., 2010). Благодаря достаточному расширению через турбину потребность в охлаждении системы LP-EGR ниже, а распределение EGR на цилиндр лучше, чем в системе HP-EGR (van Aken et al., 2007; Torregrosa et al., 2006). ). Кроме того, LP-EGR еще раз охлаждается (по сравнению с HP-EGR) через промежуточный охладитель, что приводит к более низким температурам в коллекторе (Jun et al., 2007). Однако потенциальная проблема с двигателем, работающим только на LP-EGR, заключается в том, что EGR всегда будет проходить через CAC, где может произойти конденсация и выпадение воды. Вода может мешать работе двигателя, если она конденсируется в больших количествах.
РИСУНОК 1 . Схема системы (A, слева) LP-EGR и (B, справа) HP-EGR.
Система HP-EGR, с другой стороны, полностью заключена в части высокого давления воздушной системы, до турбины и после компрессора (рис. 1B). По мере увеличения HP-EGR противодавление выхлопных газов уменьшается. Эта система основана на положительном градиенте давления для направления EGR к впускному коллектору. Во время стехиометрической работы, когда впуск дросселируется, давление во впускном коллекторе может быть ниже, чем давление выхлопа, создавая таким образом эффект вакуума, который заставляет EGR течь без помощи дополнительного противодавления. Система HP-EGR более широко используется в дизельных двигателях из-за более простой компоновки, меньшего эффекта загрязнения компрессора и лучшей реакции EGR, чем другие системы EGR. С увеличением скорости HP-EGR в условиях частичной нагрузки противодавление выхлопных газов уменьшается, что приводит к уменьшению разницы давлений между впуском и выпуском. Это помогает снизить насосные потери, связанные с уменьшением расхода топлива (Williams et al., 2017; van Aken et al., 2007).
Юго-Западный научно-исследовательский институт (SwRI) постоянно исследует новые концепции технологии двигателей в рамках нескольких внутренних исследовательских программ и Консорциума высокоэффективных бензиновых двигателей (HEDGE™). Предыдущие исследования HEDGE-II показали, что попытки согласования одного турбокомпрессора или нагнетателя с двигателем типа HEDGE оказались трудными (Joo et al., 2012). Когда оборудование повышения давления было настроено для работы на низкой скорости, диапазон расхода ограничивает работу на высокой скорости. И наоборот, при работе на высоких скоростях системы наддува были неспособны обеспечить высокие отношения давления, необходимые для работы на низких скоростях. Было высказано предположение, что двигатель HEDGE с системой HP-EGR и LP-EGR может соответствовать требованиям кривой крутящего момента с одним турбокомпрессором. Одно из предыдущих исследований (Alger et al., 2005) было проведено на бензиновом двигателе с различными степенями сжатия и конфигурациями EGR при различных скоростях и режимах нагрузки: HP-EGR с 17,5:1 CR, LP-EGR и HP-EGR. Системы рециркуляции отработавших газов с внешним наддувом при CR 12,5: 1. Исследование показало, что концепция HEDGE имеет потенциал для достижения почти таких же показателей экономии топлива, как дизельный двигатель, оборудованный для соответствия уровням выбросов MY2002/2004. Другое исследование (Mao et al., 2015), проведенное на дизельном двигателе большой мощности, показало, что двухконтурная система рециркуляции отработавших газов способна обеспечить наилучшую тепловую эффективность тормозов (BTE) и выбросы. Это произошло потому, что система рециркуляции отработавших газов с двойным контуром работала с оптимальным компромиссом между общим тепловым КПД и насосными потерями, а самые низкие выбросы были связаны с надлежащей задержкой воспламенения и коэффициентом эквивалентности. Во время этого исследования система HP-EGR могла работать наилучшим образом, когда уровень EGR был ниже 22,5%, следовательно, для системы EGR с двойным контуром контур LP-EGR включался, когда уровень EGR превышал 22,5%, а уровень LP — Доля EGR увеличилась с увеличением требуемой EGR.
При работе двигателя с системой HP-EGR было замечено наличие значительного обратного потока через контур HP-EGR. В результате этого обратного потока чистый EGR, подаваемый в цилиндры, был ниже заданного. Чтобы решить эту проблему, в систему HP-EGR был встроен односторонний клапан, который помог предотвратить этот обратный поток и привел к более высокой чистой доле EGR при сохранении разумных положений привода. В этом исследовании задокументированы подходы, использованные при настройке базового двигателя для работы со смешанными системами EGR (HP-EGR и LP-EGR), а также всесторонняя оценка использования одностороннего клапана для предотвращения обратного потока, возникающего во время работы HP-EGR.
Подход
Технические характеристики двигателя и условия эксплуатации
В этом проекте для демонстрации характеристик смешанной системы рециркуляции отработавших газов использовался 2,0-литровый двигатель KIA G4KH для легких условий эксплуатации. Технические характеристики двигателя приведены ниже в Таблице 1. Максимальная нагрузка серийного двигателя G4KH составляет 23 бар BMEP и 205 кВт.
ТАБЛИЦА 1 . Характеристики двигателя.
Уровень удельной мощности этого двигателя находится в диапазоне от среднего до высокого, однако поддержание такой высокой производительности с рециркуляцией отработавших газов было маловероятным. Следовательно, цель состояла в том, чтобы поддерживать постоянное BMEP 20 бар в диапазоне от 2000 до 5000 об/мин и производить 85 кВт/л при 6000 об/мин (достигается при BMEP 17 бар). Предлагаемая скорость рециркуляции отработавших газов 25% будет работать со стехиометрической заправкой во всех точках, но допускается снижение рециркуляции отработавших газов для достижения целевого показателя BMEP и стабилизации процесса сгорания. Один турбонагнетатель был адаптирован для удовлетворения требований к крутящему моменту на низких скоростях при работе с LP-EGR, чтобы сохранить возможности продувки. Предполагалось, что на высоких скоростях тот же турбокомпрессор сможет удовлетворить требования к крутящему моменту на высоких скоростях при работе с HP-EGR. На рис. 2 показана базовая линия и предлагаемая кривая крутящего момента.
РИСУНОК 2 . Базовые и предлагаемые кривые производительности.
Установка двигателя и базовые испытания
Двигатель был установлен вместе с блоком управления двигателем (ECU), предоставленным Hyundai Motor Company (HMC), который был загружен производственной калибровкой для проведения всех операций обкатки двигателя и для базовых испытаний двигатель. Двигатель был проверен на соответствие ожидаемому удельному расходу топлива при торможении (BSFC) в более чем 50 различных рабочих точках. Также была проверена кривая крутящего момента до 6000 об/мин. В качестве основы для будущих сравнений BSFC использовались восемь конкретных рабочих условий скорости нагрузки (показаны в таблице 2).
ТАБЛИЦА 2 . Измеренный BSFC в условиях теста HEDGE.
После проверки двигателя на испытательном стенде модель GT-Power, отправленная HMC, была проверена в соответствии с результатами двигателя. Затем модель была модифицирована, чтобы включить систему HP-EGR с набором клапанов горячей и холодной сторон, систему LP-EGR и подходящий турбонагнетатель. Первоначально турбокомпрессор OEM был масштабирован в модели, чтобы показать, что он может соответствовать целевым кривым выступа с конфигурацией Blended EGR. Затем эти результаты были использованы для подбора возможных турбокомпрессоров к двигателю. В дальнейшем в исследовании использовался увеличенный турбокомпрессор GT2056, предоставленный Garrett Turbochargers (ранее Honeywell).
Все полученные результаты были получены при целевом режиме рециркуляции отработавших газов 25 %, за исключением 2 000 об/мин и ниже, где скорость рециркуляции отработавших газов была снижена до 15 % для достижения целевого значения BMEP. Номинальная мощность 90 кВт/л (по сравнению с предлагаемой целью 85 кВт/л) была достигнута при номинальной мощности 6 000 об/мин, 18 бар BMEP и 25 % рециркуляции отработавших газов.
На рис. 3 показано, как две системы рециркуляции отработавших газов объединяются для удовлетворения общих требований к системе рециркуляции отработавших газов двигателя при полной нагрузке. При 2000 об/мин EGR обеспечивалась исключительно системой LP-EGR. Поскольку давление на входе в турбину увеличилось и потребовалось перепускание выхлопных газов через перепускной клапан, был использован HP-EGR. Общая стратегия управления EGR заключалась в том, чтобы использовать как можно больше HP-EGR при открытом перепускном клапане (рис. 4). Это связано с тем, что HP-EGR имеет меньшие насосные потери, связанные с его работой, поскольку он использует перепад давления для привода EGR.
РИСУНОК 3 . Смешанный EGR из систем HP-EGR и LP-EGR.
РИСУНОК 4 . Диаметр вестгейта при полной нагрузке.
Особое внимание уделялось характеристикам компрессора, чтобы определить, приводит ли стратегия рециркуляции отработавших газов к нежелательным рабочим областям, а именно к дросселированию или помпажу. Помпаж наиболее вероятен при низкой скорости и более высокой нагрузке, когда массовый расход низок, но степень повышения давления высока. Дроссель возникает при более высоких массовых расходах, когда компрессор достигает физического предела массового расхода. На Рис. 5 (СЛЕВА) показана смешанная рабочая область рециркуляции отработавших газов на базовом (стандартном) компрессоре (данные являются собственностью компании, поэтому значения степени сжатия, скорости, массового расхода и эффективности скрыты). Ключевым выводом из рисунка является то, что смешанная работа рециркуляции отработавших газов с базовым компрессором привела его к нежелательным рабочим областям дросселирования и помпажа, поэтому для этого исследования использовался другой компрессор (масштабированный по массовому расходу).
РИСУНОК 5 . Смешанные рабочие точки системы рециркуляции отработавших газов на карте (СЛЕВА) Базовый/стандартный компрессор и (СПРАВА) Карта эффективности нового компрессора.
Стратегия смешанной системы рециркуляции отработавших газов позволяла одному турбонагнетателю работать в широком диапазоне скоростей. На высоких скоростях HP-EGR уменьшал массовый поток через турбонагнетатель, позволяя использовать турбонагнетатель меньшего размера по сравнению с использованием LP-EGR на высоких скоростях. Стратегия LP-EGR на низких скоростях увеличила массовый поток через турбонагнетатель по сравнению с HP-EGR. Более высокий массовый расход вызывал большую работу расширения в турбине, что приводило к более низкому наддуву и смещению рабочей точки компрессора вправо (в сторону от помпажа). На рис. 5 (справа) показаны рабочие точки нового компрессора (масштабированная модель Garrett Turbochargers), указывающие на то, что поток не попадает в области помпажа и дросселирования, как это было с базовым компрессором. Этот запас будет сохранен, если EGR будет заменен на свежий воздух, чтобы работать с более высоким BMEP.
Преобразование смешанной системы рециркуляции отработавших газов
После проведения базовых испытаний и проверки модели были предприняты шаги по изменению конфигурации для реализации стратегии смешанной системы рециркуляции отработавших газов. Начальная часть этапа включала установку контуров LP-EGR и HP-EGR (с клапанами и охладителями). Система HP-EGR состояла из клапанов горячей и холодной сторон, охладителей EGR и одностороннего клапана. Оба набора клапанов были независимо оценены при моделировании и на двигателе, и было обнаружено, что при наличии клапанов горячей стороны через турбину проходит более высокий массовый расход, что приводит к большему наддуву и более высоким нагрузкам, достигаемым во время работы LP-EGR. . Из-за этого повышения производительности клапаны горячей стороны были выбраны в конфигурации HP-EGR. Требования к системе зажигания увеличились с увеличением разбавления из-за EGR. Таким образом, стандартная система зажигания была заменена прототипом системы зажигания с двойной катушкой (DCI) BorgWarner, в которой используется технология смещения двойной катушки (DCO™), ранее разработанная SwRI (Alger et al., 2011). На рис. 6 показана полная схема смешанной конфигурации EGR.
РИСУНОК 6 . Смешанная конфигурация EGR.
Производственный контроллер, использовавшийся для начальной обкатки и проверки, не был способен управлять двойными контурами рециркуляции отработавших газов, поэтому двигатель был переведен на специальный контроллер с полными полномочиями на основе cRIO (Compact Reconfigurable Input Output) от National Instruments. cRIO был сконфигурирован для работы EGR, впускных дроссельных клапанов и системы DCI. Впускной ограничительный клапан использовался для привода LP-EGR на низких скоростях, когда давление впуска было выше, чем давление выхлопа. При установке смешанных компонентов EGR была выполнена проверка контроллера SwRI. Результаты этого тестирования сравнивались с данными, собранными с помощью базового/базового контроллера (Таблица 2), чтобы гарантировать, что характеристики двигателя могут быть воспроизведены в восьми точках HEDGE. Полученная разница между штатным контроллером и контроллером SwRI, как видно на рисунке 7, показывает, что характеристики двигателя соответствуют ожидаемым допускам.
РИСУНОК 7 . BSFC Процентная разница между базовым (базовым) контроллером и контроллером SwRI.
За проверкой контроллера SwRI последовало дальнейшее моделирование и испытания двигателя для полной смешанной системы EGR. Во время испытаний было замечено, что имеется достаточный обратный поток всасываемого топлива через систему HP-EGR. В некоторых рабочих точках этот обратный поток не позволял рециркуляции отработавших газов поступать в цилиндр. В следующем разделе подробно рассказывается об использовании одностороннего клапана для предотвращения этого обратного потока.
Односторонний клапан
Как указывалось ранее, во время работы HP-EGR мгновенные пульсации давления будут возвращать всасываемый заряд обратно через контур HP-EGR. Этот обратный поток создавал более высокие пиковые значения массового расхода через контур HP-EGR и ограничивал максимальное количество HP-EGR. В некоторых рабочих условиях перепад давления между впускным и выпускным коллектором не позволял потоку HP-EGR, и главный дроссель должен был быть закрыт, чтобы создать перепад давления для приведения в действие HP-EGR. Односторонний клапан уменьшал обратный поток в контур HP-EGR и позволял использовать HP-EGR там, где перепад давления в коллекторе на двигателе не приводил к естественному приводу HP-EGR. Первоначальное моделирование показало, что существуют некоторые условия, при которых возможен обратный поток (рис. 8). Обратный поток присутствовал, когда прогнозируемый массовый расход был ниже нуля.
РИСУНОК 8 . Массовый расход HP-EGR, указывающий на противоток.
Односторонний клапан, расположенный на соединении контура HP-EGR с впускным коллектором, предотвращает этот обратный поток. Устранение обратного потока должно обеспечить больший поток HP-EGR для данной разницы давлений. Односторонние клапаны были смоделированы в GT-Power как отверстия с коэффициентом обратного нагнетания, равным нулю (рис. 9). Эта модель представляла собой максимальный потенциал одностороннего клапана.
РИСУНОК 9 . Односторонний клапан, смоделированный в GT-Power.
При 3000 об/мин, BMEP 13 бар и рециркуляции отработавших газов 15% более высокий пиковый массовый расход развивался через контур высокого давления-рециркуляции отработавших газов без одностороннего клапана (обратного клапана), как показано на рис. при низкой нагрузке и высокой скорости, а также в условиях высокой нагрузки (рис. 11). Значительный обратный поток во всех точках предполагал потенциальное расширение работы HP-EGR.
РИСУНОК 10 . Обратный поток при 3000 об/мин, 13 бар BMEP и 15 % EGR.
РИСУНОК 11 . Сравнение одностороннего клапана на (A, слева) 2000 об/мин, 2 бар BMEP; (B, справа) 4 500 об/мин, 18 бар BMEP.
Использование одностороннего клапана в контуре HP-EGR помогло снизить пиковый массовый расход примерно на 50% при 2000 об/мин, 2 бара BMEP и примерно на 25 % при 4500 об/мин, 18 бар BMEP.
При более внимательном рассмотрении графиков Log P и Log V (рис. 12) для вышеуказанных условий видно, что с самого начала нет существенной разницы между случаями с обратным клапаном и без него. Следует отметить, что это исследование проводилось в основном в одинаковых условиях между случаями, и, следовательно, на графиках Log P и Log V не наблюдалось существенной разницы.
РИСУНОК 12 . Сравнение Log P и Log V с обратным клапаном и без него на (A) 3000 об/мин, 13 бар BMEP; (B) 2 000 об/мин, 2 бар BMEP и (C) 4 500 об/мин, 18 бар BMEP.
Моделирование показало более разумное положение клапана EGR, когда система HP-EGR работает с односторонним клапаном, а не без одностороннего клапана. Это в значительной степени проявляется в условиях 3000 об/мин, 13 бар, где без одностороннего клапана более высокое положение привода на 67% обеспечивает более низкую долю рециркуляции отработавших газов в цилиндре по сравнению со случаем с односторонним клапаном (таблица 3).
ТАБЛИЦА 3 . Положение привода рециркуляции отработавших газов и рециркуляция отработавших газов в цилиндре при 3000 об/мин и 13 бар BMEP.
Наибольшее снижение пикового массового расхода с односторонним клапаном и наибольшее изменение положения привода клапана рециркуляции отработавших газов наблюдались при рабочих условиях BMEP 13 бар при 3 000 об/мин. Это существенное снижение пикового массового расхода предполагает более высокую производительность системы рециркуляции отработавших газов в данных условиях. Следовательно, это рабочее состояние было выбрано для дальнейшего изучения. GT-Power и данные двигателя указывали, что главный дроссель должен быть закрыт, чтобы приводить в действие HP-EGR в этом состоянии (шаг 1 таблицы 4). Был введен односторонний клапан (на шаге 2 в таблице 4) при сохранении того же угла дроссельной заслонки, и моделирование предсказало 8-процентное увеличение скорости HP-EGR, ограниченное только способностью турбонагнетателей производить достаточно наддува для поддержания нагрузки. Слегка приоткрытая дроссельная заслонка не идеальна для работы, но в некоторых условиях необходима для работы HP-EGR. Кроме того, клапан HP-EGR не был полностью открыт на шаге 2, что предполагало возможность дальнейшего открытия дроссельной заслонки. Шаг 3 таблицы 4 поддерживал то же количество рециркуляции отработавших газов, что и шаг 2, но теперь дроссельная заслонка была полностью открыта. Таким образом, полезная область HP-EGR может быть расширена, что приведет к возможности большего количества HP-EGR в этом состоянии. Для шага 4 в таблице 4 максимальное количество рециркуляции отработавших газов было достигнуто на уровне 27%, что снова ограничивается наддувом. Односторонний клапан смог почти удвоить скорость HP-EGR без увеличения противодавления или PMEP. Преимущество PMEP может быть лучше реализовано вблизи границы работы HP-EGR и обычно достигается за счет увеличения скорости EGR и работы WOT, обеспечиваемой односторонним клапаном.
ТАБЛИЦА 4 . Расширение работы HP-EGR с односторонним клапаном.
Результаты одностороннего клапана на двигателе были столь же многообещающими. В условиях BMEP 13 бар при 3000 об/мин они предусмотрели более разумные положения привода с точки зрения дроссельной заслонки и клапанов EGR (таблица 5).
ТАБЛИЦА 5 . Положения привода при 3000 об/мин/13 бар в условиях BMEP.
HP-EGR с односторонним клапаном также был протестирован при 1500 об/мин и в условиях высокой нагрузки. На рис. 13 показано, что без одностороннего клапана из-за обратного потока выхлопных газов в систему HP-EGR чистый поток EGR в двигатель равен 0%. Однако использование одностороннего клапана даже в условиях низкой скорости и высокой нагрузки, когда поток HP-EGR не ожидается, обеспечивает 16% чистого потока EGR. Это продемонстрировало способность одностороннего клапана расширить использование HP-EGR.
РИСУНОК 13 . Поток EGR с односторонним клапаном на низкой скорости и в условиях высокой нагрузки.
При более низком пиковом массовом расходе и более высоком уровне рециркуляции отработавших газов во время работы системы рециркуляции отработавших газов высокого давления с односторонним клапаном на турбину подается более высокое давление. Следовательно, турбина меньшего размера может использоваться для извлечения такого же количества наддува с помощью одностороннего клапана. Меньшая турбина также была бы полезна в переходных режимах, поскольку она могла бы обеспечить более быстрый отклик, что привело бы к меньшей турбо-задержке.
Заключение
В этой работе было проанализировано влияние одностороннего клапана в системе HP-EGR. При исследовании смешанной системы EGR на легком бензиновом двигателе было замечено, что во время работы HP-EGR мгновенные пульсации давления возвращали EGR обратно через контур EGR высокого давления. Для достижения желаемого уровня рециркуляции отработавших газов при заданных условиях двигатель должен был затрачивать энергию в виде насосной работы. Различные симуляции с односторонним клапаном на GT-Power показали его преимущество в предотвращении любого обратного потока через контур EGR высокого давления, что, в свою очередь, привело к расширению рабочего диапазона HP-EGR. Во время моделирования это означало гораздо более широкое положение дроссельной заслонки, что сводило к минимуму работу насоса и увеличивало пропускную способность системы HP-EGR.
Заметив многообещающие результаты моделирования, установка была реализована на движке. Результаты испытаний двигателя показали разумное положение привода с точки зрения клапана рециркуляции отработавших газов и угла дроссельной заслонки. Более широкий дроссель означал, что работа по перекачиванию значительно сократилась. Несколько лучший BSFC также наблюдался при наличии односторонних клапанов, чего не наблюдалось во время моделирования. Кроме того, способность одностороннего клапана увеличивать количество рециркуляции отработавших газов при любых условиях приводила к увеличению давления в турбине. Таким образом, турбина меньшего размера может использоваться для создания такого же наддува, чтобы соответствовать требованиям нагрузки.
Будущая работа
В этом документе исследуется влияние одностороннего обратного клапана на обратный поток через систему HP-EGR. Как уже отмечалось, во время работы HP-EGR из-за значительного обратного потока через контур HP-EGR через систему проходит более высокий пиковый массовый расход для достижения заданной скорости EGR в цилиндре. С введением одностороннего клапана пиковый поток снижается, чтобы обеспечить чистую скорость рециркуляции отработавших газов в цилиндрах. Также наблюдается то, что из-за меньшего обратного потока может быть достигнуто более разумное положение привода. Объем работы был ограничен предварительным использованием одностороннего клапана на управляемость EGR в системе HP-EGR и проверкой жизнеспособности концепции. Будущие публикации будут включать всестороннее исследование улучшений производительности в отношении BSFC и оптимизированных насосных потерь при работе смешанной системы EGR в течение нормативных ездовых циклов.
Заявление о доступности данных
Наборы данных, представленные в этой статье, были одобрены членами HEDGE. Дополнительные наборы данных являются собственностью и не всегда доступны. Запросы на доступ к наборам данных следует направлять доктору Грэму Конвею [[email protected]].
Вклад авторов
Компания BD участвовала в моделировании GT-Power смешанной системы рециркуляции отработавших газов. GC и BD провели эксперименты с двигателем, а SR внесла свой вклад в интеграцию блока управления для испытаний двигателя. GH участвовал в сборе результатов для литературы.
Финансирование
Эта работа финансировалась в рамках консорциума HEDGE-III TM Юго-Западного научно-исследовательского института.
Конфликт интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Ссылки
Алджер Т., Гингрич Дж., Мангольд Б. и Робертс К. (2011). Система зажигания с непрерывным выпуском для расширения предела рециркуляции отработавших газов в двигателях SI. Международный SAE. Дж. Инж. 4 (1), 677–692. doi:10.4271/2011-01-0661
Полный текст CrossRef | Google Scholar
Алджер Т., Ханхе С., Робертс К. и Райан Т. (2005). Бензиновый двигатель для тяжелых условий эксплуатации — многоцилиндровый образец высокоэффективной технологии с низким уровнем выбросов. Технический документ SAE 2005-01-1135. doi:10.4271/2005-01-1135
Полный текст CrossRef | Google Scholar
Джу, С., Алджер, Т., Чадвелл, К., и Де Охеда, В. (2012). Высокоэффективный бензиновый двигатель для тяжелых условий эксплуатации. Международный SAE. Дж. Инж. 5 (4), 1768–1789. doi:10.4271/2012-01-1979
CrossRef Полный текст | Google Scholar
Джун Дж., Сонг С., Чун К. и Ли К. (2007). Сравнение уровня NOx и BSFC для систем HP-EGR и LP-EGR дизельного двигателя большой мощности. Технический документ SAE 2007-01-3451. doi:10.4271/2007-01-3451
Полный текст CrossRef | Google Scholar
Maiboom, A., Tauzia, X., Shah, S., and Hétet, J. (2010). Экспериментальное исследование системы рециркуляции отработавших газов низкого давления на автомобильном дизельном двигателе по сравнению с системой рециркуляции отработавших газов высокого давления в отношении выбросов твердых частиц и NOx, а также удельного расхода топлива. Международный SAE. Дж. Инж. 2 (2), 597–610. doi:10.4271/2009-24-0138
Полный текст CrossRef | Google Scholar
Мао Б., Яо М., Чжэн З. и Ли Ю. (2015). Влияние двухконтурной системы рециркуляции отработавших газов на производительность и выбросы дизельного двигателя, Технический документ SAE 2015-01-0873. doi:10.4271/2015-01-0873
CrossRef Полный текст | Google Scholar
Парк Ю. и Бэ К. (2014). Экспериментальное исследование влияния соотношения EGR высокого/низкого давления в дизельном двигателе легкового автомобиля. Заявл. Энергия 133, 308-316. doi:10.1016/j.apenergy.2014.08.003
CrossRef Полный текст | Google Scholar
Торрегроса А., Олмеда П., Мартин Дж. и Дегреуве Б. (2006). Эксперименты по влиянию наддува и температуры охлаждающей жидкости на мощность и выбросы дизельного двигателя с прямым впрыском. Экспл. Терм. Науки о жидкости. 30 (7), 633–641.
Полнотекстовая перекрестная ссылка | Google Scholar
ван Акен М., Виллемс Ф.